



原核生物遺伝研究室・仁木研究室
比較ゲノム解析研究室・藤山研究室
生物遺伝資源情報研究室・小原研究室
大腸菌は通常は桿菌(かんきん)と呼ばれる形態をしています(図A左)。このような形態を作るためには、細胞壁、特にペプチドグリカンという堅い構造が正しく合成されなければなりません。抗生物質には、この合成を阻害し、バクテリアを殺すものがあります。しかし、これに対して抵抗性を示すバクテリアが近年増加しており、そのためもペプチドグリカンの合成をもっと理解する必要があります。
私たちは大腸菌の桿菌形態の維持に必要なRodZタンパク質を発見し、これまでその機能を研究してきました。rodZ遺伝子を破壊した株は生きられるものの生育が遅くなります。また、その形態は球形になってしまいます(図A中央)。私たちは、生育の遅いこのrodZ欠損株から、元の生育に回復した株を29株単離しました。これらは、生育が回復しただけではなく、形態も元の桿菌に戻っていました(図A右)。rodZ遺伝子は完全に破壊されているので、その機能を補うような突然変異が二次的に自然に起こったものと考えられます。これは、抑圧変異とよばれ、rodZ遺伝子の機能と関連する遺伝子を知る手がかりとなります。そこで、次世代シークエンサーを用いて、これら抑制変異株すべての全ゲノムの配列を解読しました。その結果、簡単に29株の抑制変異部位を決定することができました。
抑制変異は、
本研究は、遺伝学研究所の比較ゲノム解析研究室、生物遺伝資源情報研究室(現、先端ゲノミクス推進センター)による最新のゲノム解読の支援を受け、共同研究として行われました。
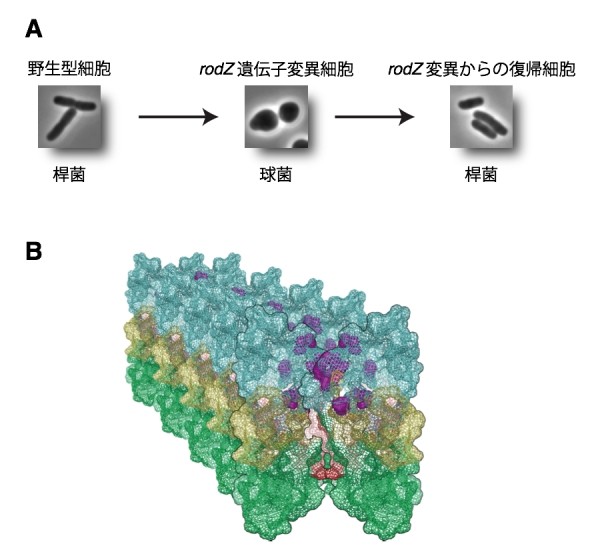
(A) 野生型の大腸菌は桿菌(左)、rodZ遺伝子変異株は球菌(中央)になります。rodZ変異からの復帰株は、元の桿菌に戻りました(右)。
(B) MreBタンパク質に見出された変異(紫色のボールでで示した)は、MreBフィラメント間の領域に見出されました。これらの変異により、MreBフィラメントの特性を変化させていると考えられます。
動原体(キネトコア)は、紡錘体微小管と結合する染色体上の重要構造体である。動原体タンパク質であるNdc80複合体は、微小管と直接結合することが知られている。しかしながら、このNdc80複合体が他の動原体タンパク質とどのように結合しているのかについては、不明な点も多い。本研究では、高解像度の構造解析によって、CENP-TのN末端側が、Ndc80複合体を構成するSpc24/25部分と直接に結合していることを明らかにした。特に、この結合はCENP-TのCDK(サイクリン依存的カイネース)によるリン酸化に依存しておこることも明らかになった。また、構造解析によって、リン酸化されたCENP-Tの72番目のT残基(T72)が直接にSpc24/25と結合するのではなく、リン酸化されたT72が近傍のリジン残基(R74)と水素結合をつくり、CENP-Tのαヘリックス構造とSpc24/25との安定な疎水結合を保証しているという分子基盤が明らかになった。さらに、生化学的な解析によって、CENP-TはNdc80複合体と結合するMis12複合体とは排他的に、Ndc80複合体と結合することも明らかになった。今回の研究により、動原体の分子構成の実体の理解が深まった。
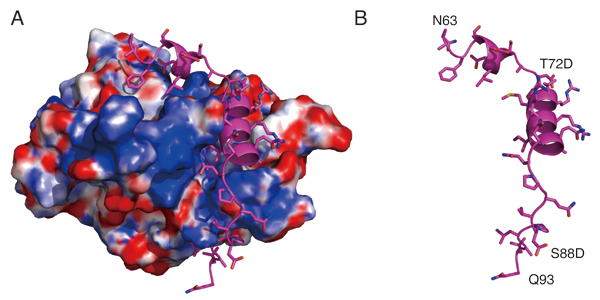
(A)Spc24/25複合体に結合したCENP-T構造(シアン)
(B)Aの図からCENP-Tのみ取り出した構造リボン図
 細胞遺伝研究部門の小林武彦教授が第29回井上学術賞を受賞しました。
細胞遺伝研究部門の小林武彦教授が第29回井上学術賞を受賞しました。哺乳類において精巣を作るか卵巣を作るかという性の決定機構は比較的よく理解されています。Y染色体にのっているSRY遺伝子が発現すると精巣が、発現しないと卵巣ができます。また生殖細胞が精子になるか卵子になるかという決定は、生殖細胞の前駆体である始原生殖細胞が精巣に入るかあるいは卵巣に入るかによって決まります。しかし、実際に生殖細胞の性分化誘導機構は理解されていませんでした。今回総研大(D5)の呉泉君が雄性生殖細胞の誘導因子を同定しました。彼はNodalシグナル系が雄性生殖細胞特異的に活性化していることを見いだし、特異的な阻害剤及びNodalシグナル系の条件付きノックアウトマウスを用いた解析を行いました。その結果、体細胞から分泌されるFGF9により生殖細胞でNodalが活性化し、その下流で雄性特異的遺伝子Nanos2が誘導されることを明らかにしました。またこの時同時に、FGFシグナルは体細胞にも機能し、主にActivinの活性化を介して体細胞の雌化を抑制することも示しました。
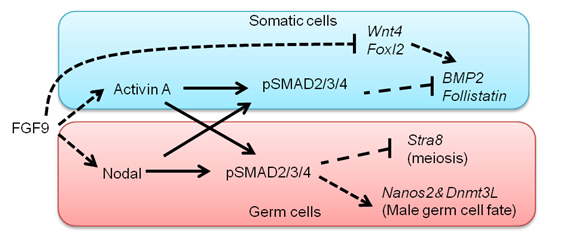
雄の性分化にTGFbシグナルであるNodal/Activinが重要な機能を果たす。FGF9の下流で体細胞ではActivinが誘導され雌化因子(BMP2、Follistain)を抑制し、体細胞の雌化を抑制する。一方、生殖細胞で特異的に活性化するnodalシグナルは生殖細胞が雌化することを抑制し、雄化遺伝子(Nanos2, Dnmt3L)の発現を誘導する。










