KNOTTED1 Cofactors, BLH12 and BLH14, Regulate Internode Patterning and Vein Anastomosis in Maize
Katsutoshi Tsuda, Maria-Jazmin Abraham-Juarez, Akiteru Maeno, Zhaobin Dong, Dale Aromdee, Robert Meeley, Toshihiko Shiroishi, Ken-ichi Nonomura and Sarah Hake.
The Plant Cell. published online April 5, 2017 DOI:10.1105/tpc.16.00967
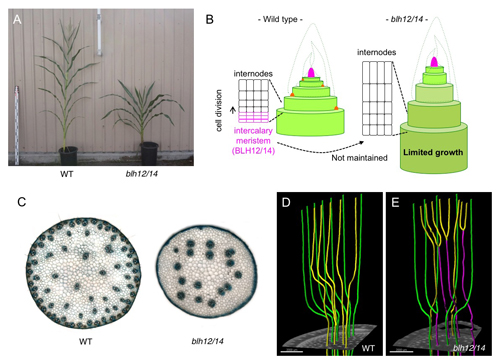
単子葉植物の茎は、維管束がリング状に並んだ双子葉植物と異なり、茎全体に散らばった散在型維管束を持ちます。また、タケにみられるように明確な節と節間の繰り返し構造からなる茎も単子葉類の特徴です。これらは理科の教科書などに出てくる良く知られた性質であり、脆弱な茎は農作物の倒伏による収量のロスをまねくため育種上重要なターゲットとされてきたのにも関わらず、その発生過程を制御する分子メカニズムはほとんど不明でした。わたしたちは、トウモロコシのBELL1-like Homeobox (BLH)転写因子がこれらの単子葉類に特徴的な茎の形態形成に重要な役割を果たしていることを明らかにしました。まず、植物の地上部全体の源である茎頂分裂組織(メリステム)の維持に不可欠なKnotted1-like Homeobox (KNOX)転写因子のコファクターとしてBLH12 およびBLH14を同定しました。blh12/14 二重変異体では、腋芽や花序のメリステムが正常に維持されないのに加え、茎が短くなる矮性を示し、その内部の維管束数は大きく減少していました。タンパク質の発現パターンの解析やマイクロCTによる茎の内部構造の詳細な観察により、BLH12/14が(1)節間基部にある介在分裂組織に蓄積し分裂活性を維持することで節間細胞の持続的な供給および十分な茎の成長を可能にしていること、(2)未熟な茎の内部で維管束同士の早期融合を抑制することで、多数の独立した維管束の形成を可能にしていることが示唆されました。本研究は国立遺伝学研究所・実験圃場の津田勝利助教とカリフォルニア大学バークレー校Sarah Hake教授を中心としたグループの共同研究の成果であり、科研費若手(B)(JP16K18637) の支援を受けて行われました。また、哺乳動物遺伝研究室の前野哲輝技術員によるマイクロCT観察がこれまで観察が困難であった維管束ネットワークの可視化において大きく貢献しました。