



RNA interference-independent reprogramming of DNA methylation in Arabidopsis
Taiko Kim To, Yuichiro Nishizawa, Soichi Inagaki, Yoshiaki Tarutani, Sayaka Tominaga, Atsushi Toyoda, Asao Fujiyama, Frédéric Berger, Tetsuji Kakutani
Nature Plants 2020 November 30 DOI:10.1038/s41477-020-00810-z
動物や植物のゲノムは、可動性で増殖する性質のDNA(transposable element: 以下TEと略する)を多量に含み、これがゲノムの不安定化や癌などの疾病の原因になります。TEを不活性化する機構として、ヒストンH3のリジン9(H3K9)のメチル化やDNAのシトシンメチル化などが知られています。このような抑制目印は、細胞分裂後にもエピジェネティックに継承、維持されます。「維持」に加え、抑制目印を「確立」するための機構として、短いRNAによるRNA干渉という機構が動物でも植物でもよく研究され、その重要性が示されてきました。
本研究では、抑制目印の「維持」に必要な因子におけるシロイヌナズナの変異体を用いて、ゲノム中のTEからこれら抑制目印が喪失した後に、抑制修飾をになう酵素を再導入することで、抑制目印の「確立」を直接調べました。その結果、驚いたことに、TE中の遺伝子コード領域(TEの増殖に必要な酵素などをコードする遺伝子領域)においては、良く知られたRNA干渉経路とは独立の経路によって、抑制目印が正確、かつ効率的に回復することがわかりました。さらに、抑制目印が回復しない少数のTEを調べることで、ヒストンH3K9meの除去に関わる酵素、ヒストンH2Aバリアント、 転写開始点付近のCG配列のメチル化、およびこれらの目印への転写からのフィードバックが抑制修飾の新規確立に重要なことが示唆されました。これらの因子は植物のみならず動物にも保存されています。本研究から明らかとなりつつある植物の抑制目印を確立する機構が、他の生物にも保存されている可能性があります。
本研究は、情報・システム研究機構 新領域融合研究プロジェクト「生命システム」として実施しました。
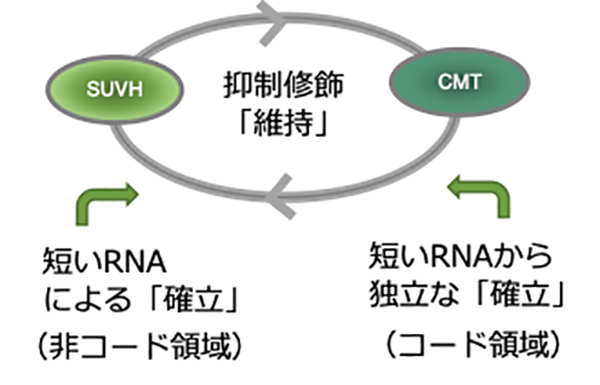
図:転移因子を抑制する機構のモデル図
SUVH(H3K9メチル化酵素)とCMT(非CG配列のDNAメチル化酵素)との相互増強によって抑制修飾が維持される。一方、抑制修飾の確立は、RNA干渉の経路に加え、これとは独立の経路がコード領域で働く。維持経路は修飾を拡張するため、RNA干渉による経路とよらない経路とが相互に補いあう。










