



Release of condensin from mitotic chromosomes requires the Ran-GTP gradient in the reorganized nucleus.
Keita Aoki and Hironori Niki
Biology Open, November 15; 6(11):1614-1628, 2017 DOI:10.1242/bio.027193.
細胞周期の分裂期に高度に凝縮した染色体は、その後の間期で脱凝縮します。染色体の脱凝縮には、凝縮因子として有名なコンデンシンが染色体から解離する必要があると考えられていますが、その機構はまだよくわかっていません。
原核生物遺伝研究室では、ヒト細胞と同様に分裂期後に核膜再生と染色体脱凝縮が協調して起こるSchizosaccharomyces japonicus(分裂酵母の一種)を真核細胞のモデルとして、この問題に取り組みました。
本研究では、まず、脱凝縮の異常変異体として染色体が過凝縮する変異株をS. japonicusの温度感受性変異株ライブラリーから見つけることにしました。見つかった過凝縮する変異株は、pim1遺伝子に変異があることがわかりました。Small GTPase Ranのグアニンヌクレオチド交換因子であるPim1/RCC1の変異株では、染色体が異常に強く凝縮することが分裂酵母やハムスターを用いた研究から予てより知られていました(Nishimoto et al., 1978; Matsumoto and Beach 1991; Sazer and Nurse 1994)。
次に、S. japonicusの染色体過凝縮表現型の原因を明らかにするために、Pim1の細胞内局在を野生型と変異型とで比べました。そうしたところ、野生型Pim1は細胞周期を通じて主に核膜孔に局在していました。一方、変異型Pim1は、核膜孔に局在しましたが、この局在は核分裂後に娘細胞のうち片方の核から消失していました。そして、Pim1局在の消失した核でのみ染色体の過凝縮が起こることを我々は見つけました。
では、なぜPim1の核膜孔局在が不安定になると染色体の過凝縮が起こるのでしょうか?我々は、原因としてコンデンシンの影響を考えました。コンデンシンの染色体局在は野生型では分裂期後になくなりますが、pim1変異株では、過凝縮した染色体にコンデンシンが残留していました。また、pim1変異株の過凝縮染色体の頻度はコンデンシンの活性化リン酸化サイトの一つであるCDKサイトの活性化型変異により増加しました。これらの結果から、Ran経路に依存したコンデンシンの脱リン酸化もしくは不活性化が、効率的な染色体脱凝縮に関わると考えられました。
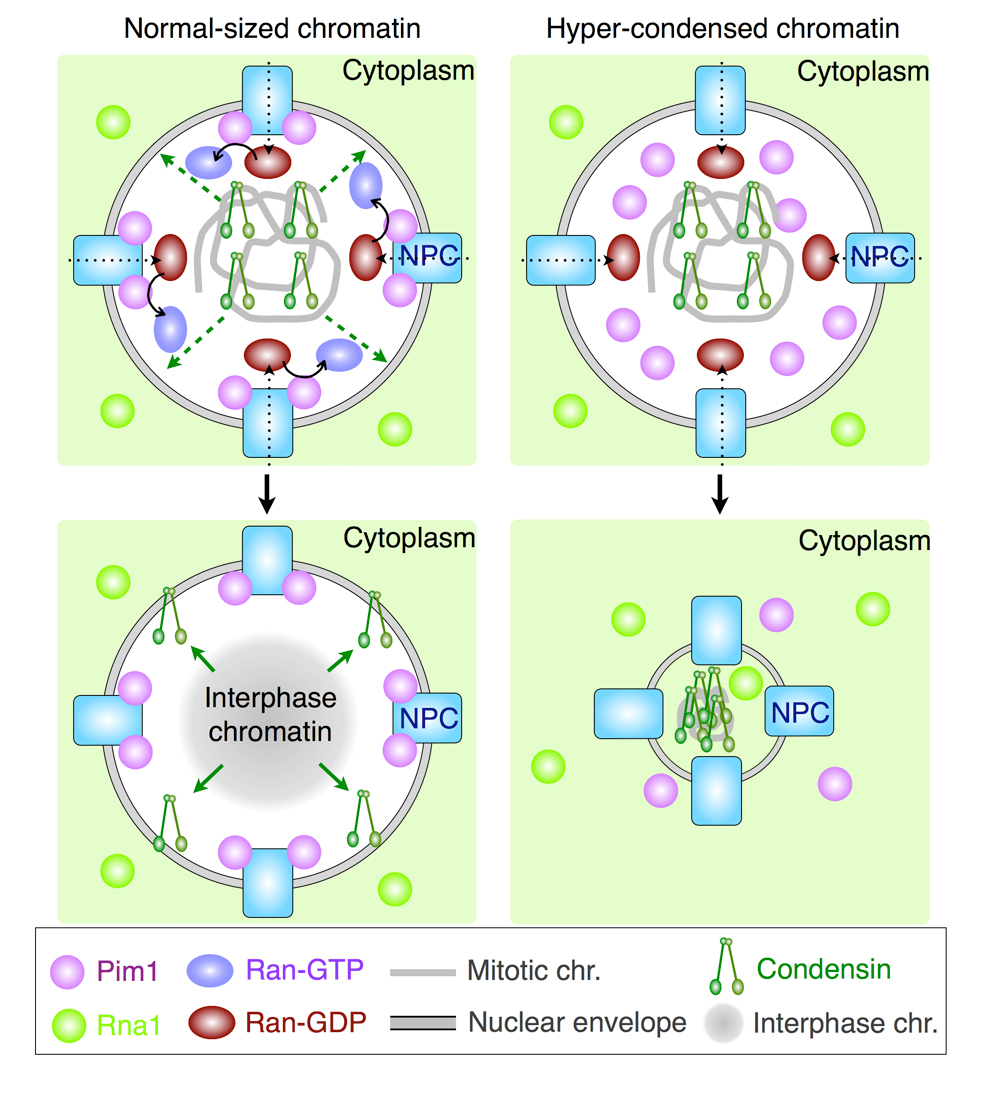
通常の核(左側)では、染色体上のコンデンシンは、分裂期後にRan-GTPに依存して染色体から離れる。そのため、脱凝縮したG1期核が形成される。一方、Pim1/RCC1に異常がある核(右側)では、Ran-GTPが少ないために分裂期後にコンデンシンが染色体から離れることができない。そのため、G1期に染色体が過凝縮する。










