



細胞運命の維持は、発生及び生物の恒常性に不可欠であり、その異常は必要な細胞の欠損や癌化を引き起こすと考えられる。以前に私達はC. elegansを用いヒストンのアセチル化が細胞運命の維持に関与する事を示している。運命維持には、MYSTファミリーヒストンアセチル化酵素MYS-1, MYS-2とアセチル化ヒストン結合蛋白質BET-1が必要だが、詳細な分子機構は不明であった。
BET-1の酵母相同分子BDF1はクロマチンリモデリング因子SWR1と複合体を形成し、ヒストンH2AバリアントHTZ1/H2A.zのゲノム上の配置を制御する事が知られている。私達はC. elegansにおいて、BET−1に加え、SSL-1/SWR1とHTZ-1/H2A.zが運命維持に必要である事を明らかにした。これらの因子は生殖巣の体細胞で細胞運命を維持する事で、生殖幹細胞のニッチ細胞であるDTCの過剰形成を抑制する。私達はDTCの誘導に必要な転写因子をコードするceh-22がHTZ-1の直接の標的遺伝子であり、BET-1, MYS-1, HTZ-1がその転写を抑制している事を示した。ヒストンアセチル化は転写の活性化に関与する事が広く知られているが、この結果はヒストンアセチル化が転写抑制も行う事を示唆している。
また、細胞運命の維持にはH3K27のメチル化が関与する事が以前から知られていた。私たちは、ceh-22遺伝子へのHTZ-1の局在はH3K27脱メチル化酵素UTX-1の阻害で減少すること、HTZ-1とH3K27メチル化のゲノム上の局在には負の相関があることを示し、HTZ-1はUTX-1によってメチル化が除かれた場合に、転写抑制を行うことで細胞運命を維持している事を示唆した。H3K27のメチル化による安定した転写抑制と異なり、BET-1やHTZ-1は、分化を促すシグナルによって転写が簡単に誘導されるような遺伝子の、シグナルに依存しない転写を抑制していると考えられる。
この研究は関西学院大学・柴田博士、西脇博士との共同研究として行なわれました。
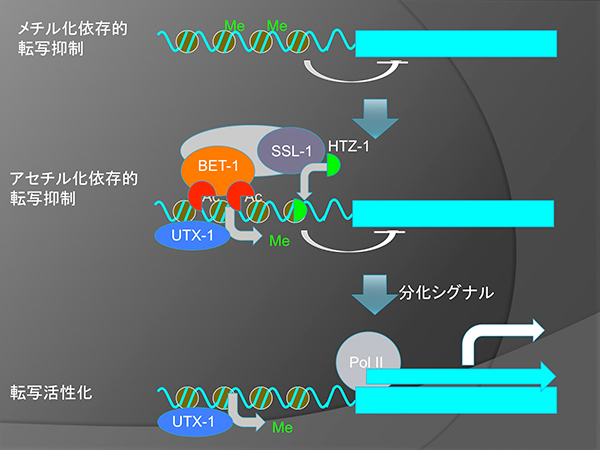
転写活性化の二段階モデル。H3K27のメチル化による安定な転写抑制がUTX-1によって解除されると、遺伝子は転写活性化可能となるが、分化シグナルを受けない場合は、BET-1とHTZ-1によって抑制されている。










