



Distinct functions of three Wnt proteins control mirror-symmetric organogenesis in the C. elegans gonad
Shuhei So, Masayo Asakawa and Hitoshi Sawa
eLife (2024) Nov 1:13:e103035 DOI:10.7554/eLife.103035
多くの器官は鏡像対称な形をしています。そのためには器官形成時に、同等な位置にある細胞が同じ運命を持ち、鏡像対称な挙動・移動をする必要があります。
線虫C. elegansの生殖巣は前後方向に鏡像対称な構造をしています。これは、生殖巣の伸長をガイドする二つのdistal tip細胞(DTC)が、それぞれ前方、後方に移動するからです。DTCはその前駆細胞(Z1とZ4)から二回の非対称分裂を経て作られます。Z1とZ4は鏡像対称な極性を持つために、DTCも極像対称な位置(生殖巣の遠位端)に作られると考えられます。
今回、多細胞構築研究室の澤斉教授、宗修平研究員(当時)らは、SGP細胞の鏡像対称な極性が三種類のWntタンパク質(シグナル分子)とWnt受容体であるLIN-17/Frizzledの冗長的働きによって制御されていることを明らかにしました。LIN-17はWnt受容体ですが、Wntと無関係に働きます。また三種類のWnt(CWN-1、CWN-2,EGL-20)は全く異なる効果を持っています。CWN-2は、Z1とZ4に反対向きの(つまり鏡像対称な)極性を、CWN-1はZ1・Z4共に同じ方向の極性を誘導します。さらにEGL-20は、CWN-1の働きを阻害します。lin-17 egl-20 cwn-2三重変異体では、Z1細胞の極性が逆転し、Z4細胞と同じ方向に極性化する結果、Z1由来のDTCは生殖巣の前端ではなく中央に作られます。そして、Z1由来のDTCは、後方に移動してしまいます。鏡像対称な極性形成により、DTCが生殖巣の遠位端に正しく配置されることで、鏡像対称な細胞移動および器官形成が制御されていることが明らかになりました。
本研究は科研費(JP16H04797)、武田科学振興財団およびNIG-JOINTの支援を受けて実施しました。
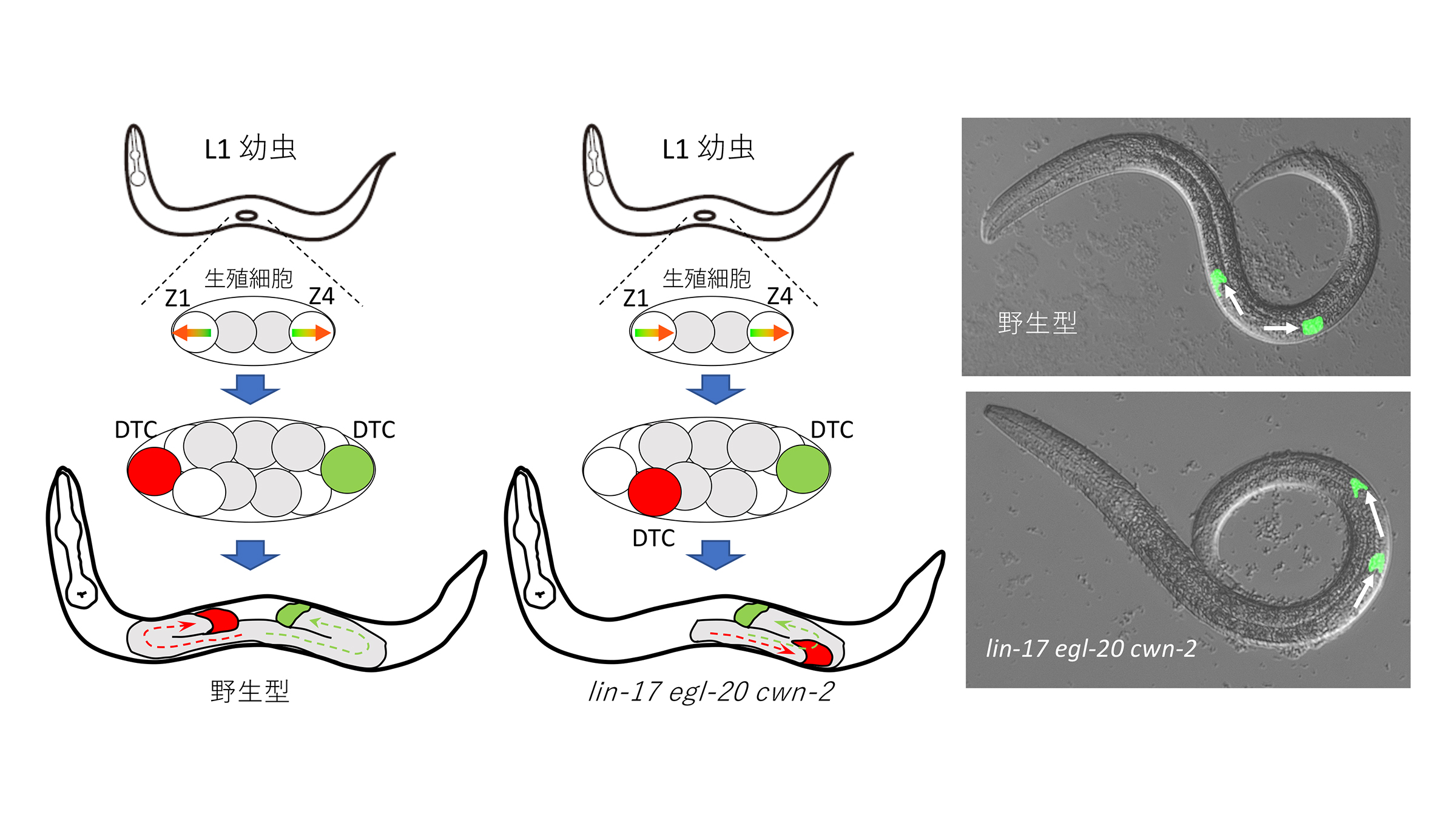
図:左:C. elegans野生型の生殖巣の発生。Z1、Z4細胞は鏡像対称な極性を持ち、二つのDTCを生殖巣の遠位端に作り出す。それぞれのDTCは遠位方向に移動する。中央;lin-17 egl-20 cwn-2変異体では、Z1の極性が逆転し、DTCが生殖巣の中央に作られ、Z1由来のDTCが後方に移動する。右:移動中のDTC(緑)の写真










