



The physical size of transcription factors is key to transcriptional regulation in chromatin domains
Kazuhiro Maeshima, Kazunari Kaizu, Sachiko Tamura, Tadasu Nozaki, Tetsuro Kokubo, and Koichi Takahashi真核生物には、大小様々な大きさの転写因子があります。そして、これらの転写因子は、微小な空間に折り畳まれている長いDNAから、必要な遺伝情報を読み出すのに必要です。一般的に、遺伝子特異的な働きをする転写因子のサイズは小さく (~50 kDa)、これらはゲノムDNA中の標的となる調節領域を探して結合します。そして、この小さい転写因子が、基本転写因子、メディエーター、RNAポリメラーゼ、ヌクレオソームリモデラー、ヒストン修飾因子など、巨大タンパク複合体 (1-3 MDa以上) の標的配列への結合を誘導すると考えられています。最近、細胞核内では、ヌクレオソーム線維が凝集することによってできるクロマチンドメイン(topologically associating domains, 図の青ボール集団) が多数形成されていることが分かってきました。私たちは、この凝集したクロマチンドメインにおける転写制御に、転写因子やその複合体の物理的なサイズが重要であるという新しいモデルを提唱しました。
私たちは、まずモンテカルロ・シミュレーションによって、凝集したクロマチンドメイン内部に入ることができる転写因子の物理的なサイズを決定しました。その結果、遺伝子特異的な転写因子群(小さいサイズ)は、ドメイン内部に入り込むことができる一方(図の黄ボール)、サイズが大きな転写複合体群は、ドメイン内部に入ることはできないことが分かりました(図の緑ボール)。
この結果、サイズの小さな転写因子がクロマチンドメイン内の標的配列を探索して結合し(図a)、その転写因子-標的配列複合体がドメイン表面に出てきた際、この転写因子を「目印」として、サイズの大きい転写因子複合体が標的配列に結合すると考えました (図b)。標的配列に結合した巨大な転写因子複合体は、ドメイン内部へと動くことが難しいため、標的配列をドメインの表面に維持する「ブイ(浮き袋)」のような働きができます(図b)。そして、ヌクレオソームリモデラー、ヒストン修飾因子など、他の巨大複合体と共に安定的な転写を可能にするという新しいモデルを提唱しました (図c)。
これらの転写因子の物理的サイズに依存した、標的探索の方法や、標的配列をドメイン表面につなぎ止める性質は、DNA複製や修復、組換えを含む様々なDNA機能に共通するメカニズムと考えられます。
本研究は理研・生命システム研究センター高橋恒一・海津一成、横浜市立大学・古久保哲朗らとの共同研究としておこなわれ、英国Journal of Physics: Condensed matter誌の「Physics of chromatin」特集号に掲載されました。
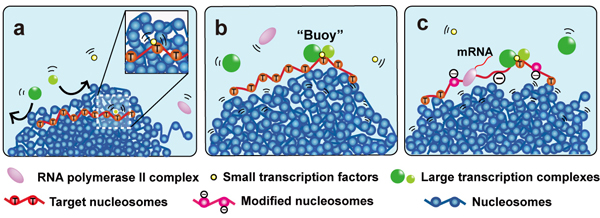
転写制御における「ブイ(浮き袋)」モデル
凝集したクロマチンドメインは青いボール集団で示している。
(a) 黄色い小さな転写因子は凝集したクロマチンドメイン内部を動くことができるが、緑の大きい転写因子複合体はドメインに入ることができない。 (b) 大きな転写因子複合体がブイ(浮き袋)となって、標的配列がドメイン表面で維持される。(c) ヌクレオソームリモデラー、ヒストン修飾因子など、他の巨大複合体と共に、安定的な転写がおこる。










