



Coordination of polyploid chromosome replication with cell size and growth in a cyanobacterium.
Ryudo Ohbayashi, Ai Nakamachi, Tetsuhiro Hatakeyama, Yu Kanesaki, Satoru Watanabe, Taku Chibazakura, Hirofumi Yoshikawa, and Shin-ya Miyagishima
mBio 10(2), e00510-19 DOI:10.1128/mBio.00510-19
Homologous chromosome number (ploidy) has diversified among bacteria, archaea, and eukaryotes over evolution. In bacteria, model organisms such as Escherichia coli possess a single chromosome. In contrast, other bacteria, including cyanobacteria, maintain multiple copies of individual chromosomes (polyploid). Although a correlation between ploidy level and cell size has been observed in bacteria and eukaryotes, it is poorly understood how replication of multi-copy chromosomes is regulated and how ploidy level is adjusted to cell size. Here we show that only one or a few multi-copy chromosomes are replicated at once in the cyanobacteria Synechococcus elongatus and that this restriction depends on regulation of DnaA activity. When cell growth rate was increased or decreased, DnaA level, DnaA activity, and the number of replicating chromosomes also increased or decreased in parallel, resulting in nearly constant chromosome copy number per unit cell volume. Thus, it is suggested that the stepwise replication of the genome enables cyanobacteria to maintain nearly constant gene copy number per unit cell volume.
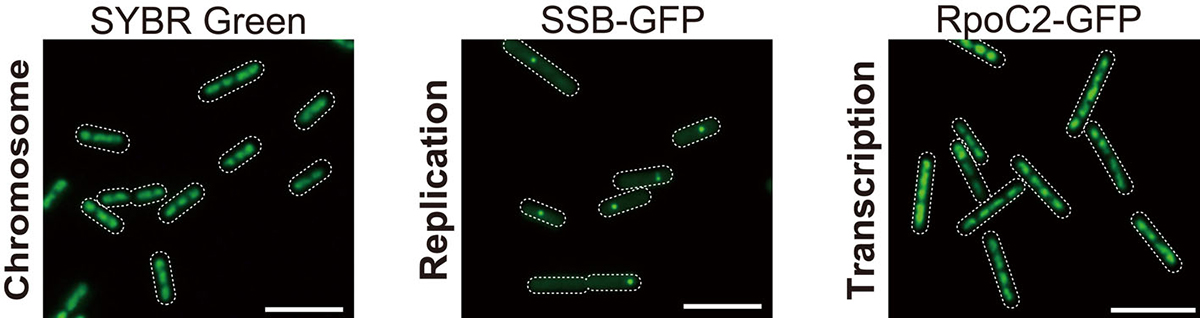
Figure. 1: Microscopic images of SYBR Green-stained nucleoids, SSB-GFP (replicating chromosomes), and RpoC2-GFP (transcribed chromosomes). All copies of chromosomes are transcribed while chromosomes are replicated one by one.
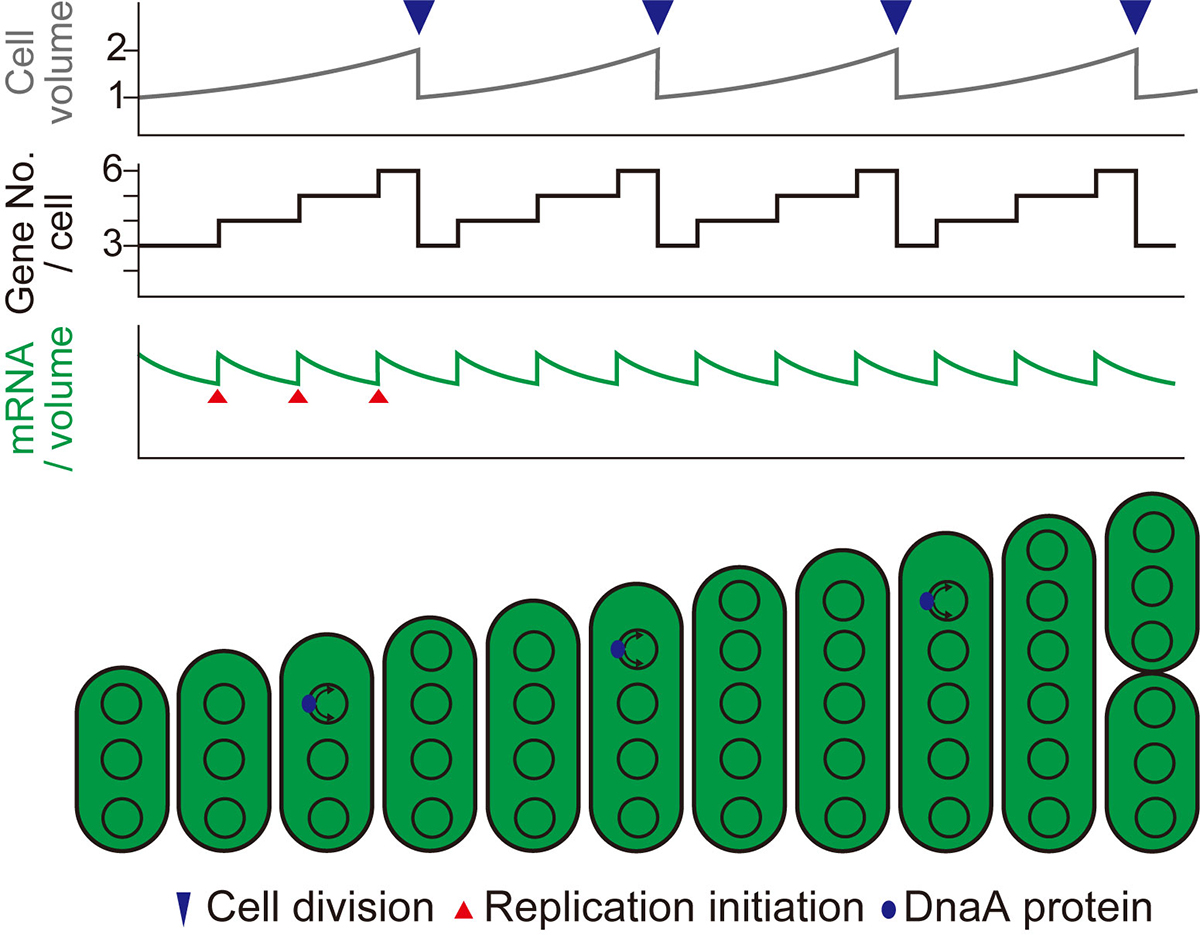
Figure. 2: Schematic diagrams showing changes in cell volume (upper), gene copy number per cell (middle), and mRNA level per unit volume (bottom) during cell growth in the case of stepwise (as observed in this study) replication of multi-copy chromosomes.
Press release
A key metabolic gene for recurrent freshwater colonization and radiation in fishes.
Asano Ishikawa, Naoki Kabeya, Koki Ikeya, Ryo Kakioka, Jennifer N. Cech, Naoki Osada, Miguel C. Leal, Jun Inoue, Manabu Kume, Atsushi Toyoda, Ayumi Tezuka, Atsushi J. Nagano, Yo Y. Yamasaki, Yuto Suzuki, Tomoyuki Kokita, Hiroshi Takahashi, Kay Lucek, David Marques, Yusuke Takehana, Kiyoshi Naruse, Seiichi Mori, Oscar Monroig, Nemiah Ladd, Carsten J. Schubert, Blake Matthews, Catherine L. Peichel, Ole Seehausen, Goro Yoshizaki, and Jun Kitano.
Science 31 May 2019: Vol. 364, Issue 6443, pp. 886-889 DOI:10.1126/science.aau5656
EurekAlert! link about this artcle
Fishes are present in not only marine but also freshwater environments including rivers, streams, lakes, and ponds because ancestral marine fishes have colonized freshwater multiple times and diversified in the freshwater environment during evolution. However, only few fish lineages could colonize freshwater habitats. What enable some lineages to colonize freshwater?
Dr. Asano Ishikawa and Dr. Jun Kitano of Ecological Genetics Laboratory at the National Institute of Genetics, Japan, collaborated with researchers from Japan, Switzerland, USA, and Spain and asked this question using stickleback fishes (Fig. 1) as a model. They identified a key gene important for freshwater colonization, Fatty acid desaturase 2 (Fads2), encoding an enzyme involved in the synthesis of docosahexaenoic acid (DHA), a polyunsaturated fatty acid. DHA is a component of the cell membrane and is essential for growth and survival of fish. In aquatic ecosystems, marine-derived diets are rich in DHA, while diets in freshwater environments comprise limited DHA. They found that sticklebacks that successfully colonized freshwater have higher copy numbers of Fads2 and higher abilities to synthesize DHA than those that failed to colonize freshwater (Fig. 2). Analysis of publicly available whole-genome sequences of 48 fish species revealed higher Fads2 copy numbers in freshwater fish than in marine fish across diverse fish taxa.
This research was supported by JSPS KAKENHI (15H02418, 23113007, 16H06279, 26870824, 16K07469), JSPS PD (11J04816, 16J06812), Asahi Glass Foundation, Sumitomo Foundation, SNF grant (31003A 175614), and SNSF (31003A_163338, PDAMP3_123135).
This was published in Science at 2:00 p.m. U.S. Eastern Time on Thursday, 30 May 2019.

Fig: 1.A photo of a Japan Sea stickleback (Gasterosteus nipponicus) male. Photo by Yasuyuki Hata.
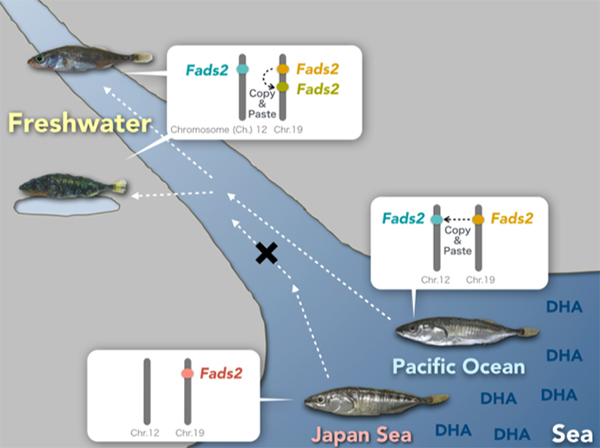
Fig: 2.Freshwater colonization and increase in Fads2 gene Japan Sea sticklebacks that could not colonize freshwater have only one Fads2 copy on Chromosome 19, while three-spined sticklebacks that could colonize have additional copy of Fads2 on Chromosome 12. Some freshwater populations of three-spined sticklebacks further duplicated Fads2 genes on Chromosome 19.
Mechanically distinct microtubule arrays determine the length and force response of the meiotic spindle
Jun Takagi, Ryota Sakamoto, Gen Shiratsuchi, Yusuke T. Maeda, Yuta Shimamoto
Developmental Cell, Vol 49, pp 267-278, 2019 DOI:10.1016/j.devcel.2019.03.014
During cell division, our genome must be faithfully transmitted from the mother cell to daughter cells. This task is done by the spindle, the micron-sized bipolar force-generating machinery. Using a microfabricated force sensor and high-resolution microscopy, a team of Yuta Shimamoto at National Institute of Genetics and Yusuke Maeda at Kyushu University probed the local mechanical architecture of the spindle (Fig. 1A) and found its substantial heterogeneity; the equator and pole regions are stiff whereas its middle is more flexible and deformable (Fig. 1B). The spindle’s mechanical heterogeneity explains how this micron-sized structure can maintain its local functional architectures, such as the pole and the equator, required to pull chromosomes and determine the cell division axis, while adapting the overall structure to perturbations. Their physical approach will help understand the mechanisms ensuring the fidelity of chromosome segregation, which is essential for normal cell proliferation and embryonic development in humans.
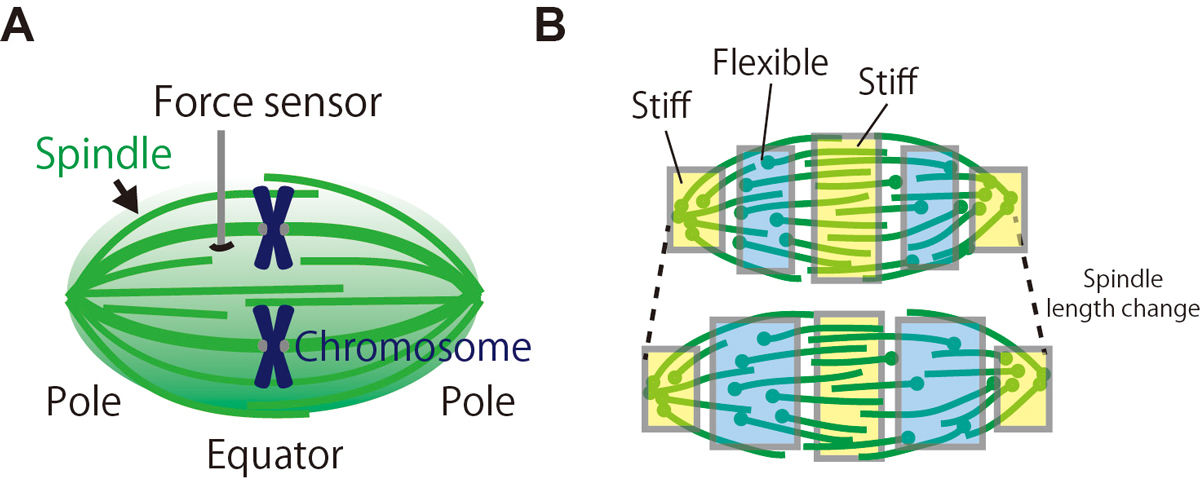
Figure: (A) The local mechanical architecture of the spindle was probed using a fiber-shaped microforce sensor. (B) The spindle was found to have a substantial mechanical heterogeneity (depicted in different colors), suggesting its robust yet adaptable nature required for error-free cell division.
Generation of conditional auxin-inducible degron (AID) cells and tight control of degron-fused proteins using the degradation inhibitor auxinole
Aisha Yesbolatova, Toyoaki Natsume, Ken-ichiro Hayashi, Masato T. Kanemaki
Methods Available online 24 April 2019 DOI:10.1016/j.ymeth.2019.04.010
The auxin-inducible degron (AID) technology developed by the Kanemaki group is based on a plant-specific degradation pathway transplanted to non-plant cells. In human cells expressing a plant E3 ligase component, TIR1, a degron-fused endogenous protein is degraded within 15–45 min upon addition of the phytohormone auxin.
We demonstrated previously the generation of human HCT116 mutants in which the C terminus of endogenous proteins was fused with the degron by CRISPR–Cas9-based knock-in (Natsume et al. Cell Reports, 2016). In this study, A SOKENDAI student, Aisha Yesbolatova, developed new plasmids for N-terminal tagging and described a detailed protocol for the generation of AID mutants of human HCT116 and DLD1 cells (Figure 1).
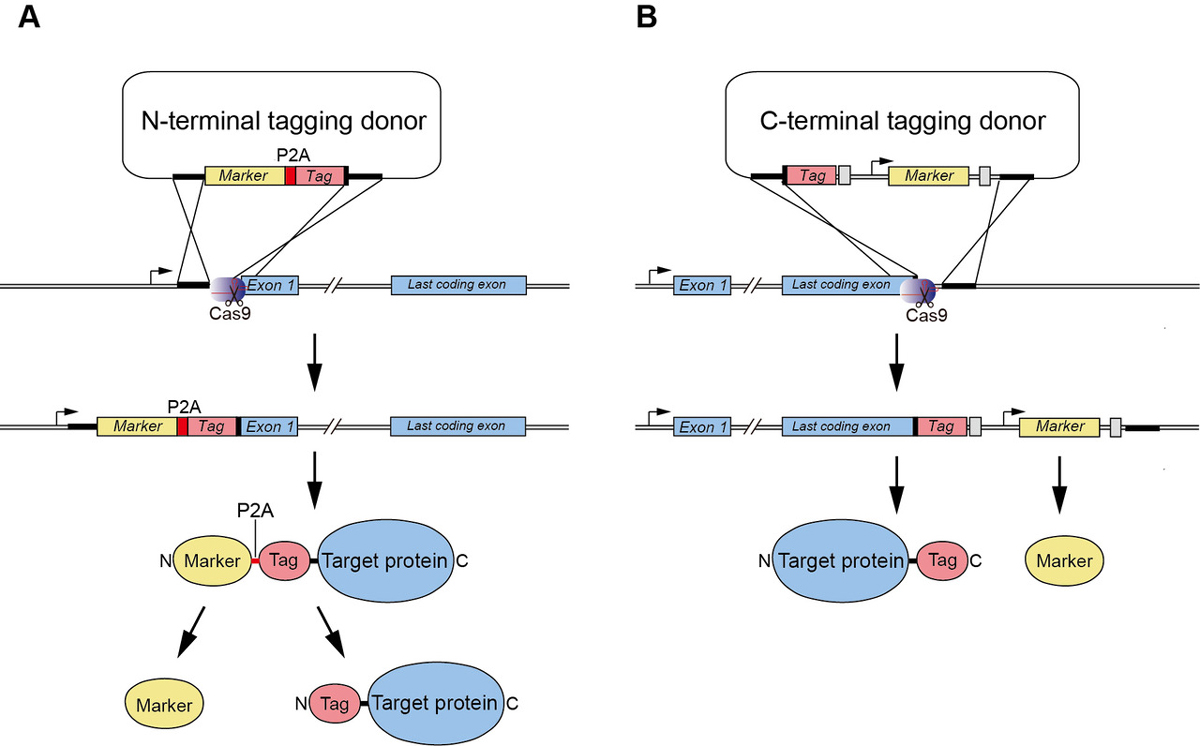
Figure1: (A) A new CRISPR/Cas9-based N-terminal tagging system. (B) C-terminal tagging.
Moreover, we report the use of a TIR1 inhibitor, auxinole, to suppress leaky degradation of degron-fused proteins (Figure 2). The addition of auxinole is also useful for rapid re-expression after depletion of degron-fused proteins.
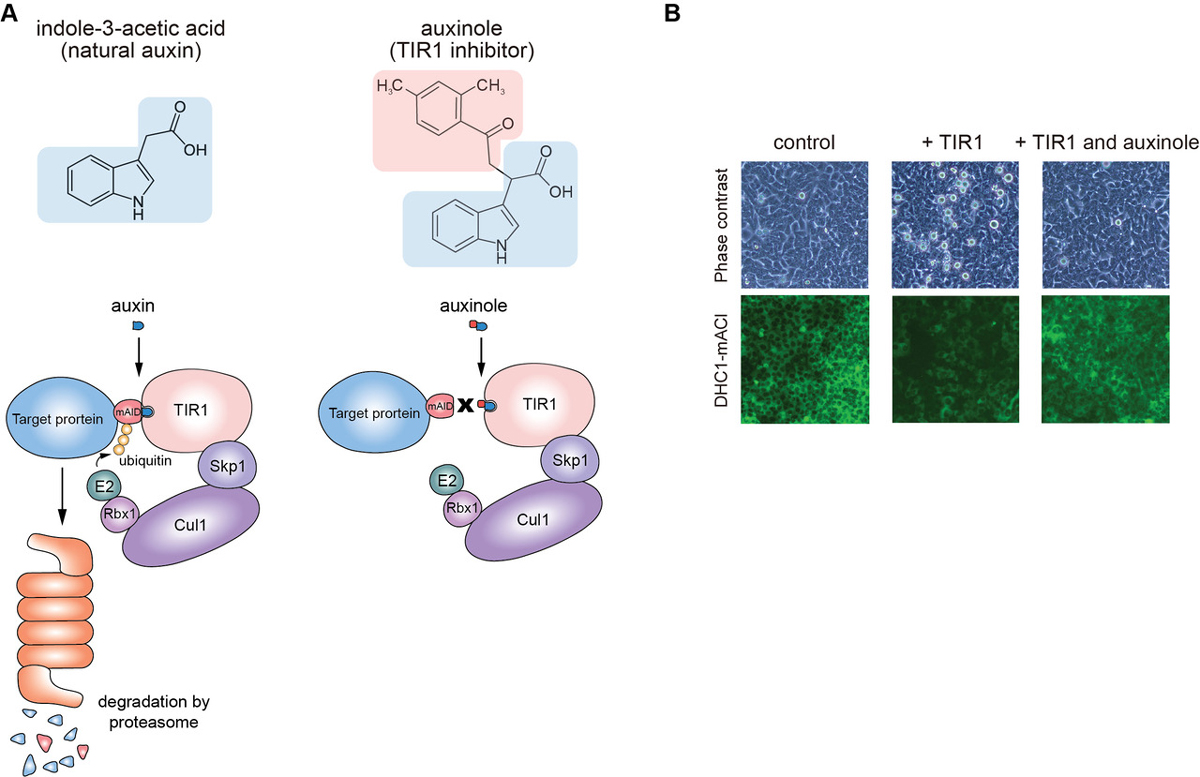
Figure2: (A) The structure of auxin and auxinole, and illustrations showing how these reagents work in the degradation pathway. (B) Suppression of leaky degradation in DHC1-tagged Tet-TIR1 cells. The cells were treated with only doxycycline to induce TIR1 expression or together with auxinole 48 h before microscopy.
We hope that these improvements enhance the utility of the AID technology for studying protein function in living human cells. All described plasmids are available from addgene and RIKEN-BRC.
This study was carried out as a collaboration with Prof. Ken-ichiro Hayashi of Okayama University of Science and was supported by JSPS KAKENHI (17K15068, 18H02170 and 18H04719), JST A-STEP (grant number AS2915150U), the Canon Foundation, the Asahi Glass Foundation, and the Takeda Science Foundation.
Frequent nonrandom shifts in the temporal sequence of developmental landmark events during teleost evolutionary diversification
Fumihiro Ito, Tomotaka Matsumoto, Tatsumi Hirata
Evolution & Development 18 April 2019 DOI:10.1111/ede.12288
Development of multicellular organisms is characterized as a series of morphological events, whose temporal sequence, called as the developmental sequence, is well conserved in each individual species. In this study, we hypothesized that evolutionary changes in the sequence can be the basis for morphological diversifications, and examined the evolutionary dynamics of the developmental sequence in teleosts. Using the information from previous reports describing the development of 30 teleost species, we extracted the developmental sequences of 19 landmark events involving the formation of phylogenetically conserved body parts. Ancestral developmental sequences were then estimated by two different parsimony based methods. The phylogenetic comparisons of these sequences revealed that (1) the frequency of sequence changes differs among the events, (2) most of the sequence changes occurred as exchanges of temporally neighboring events, and (3) these changes in developmental sequences accumulate along the evolutionary time.
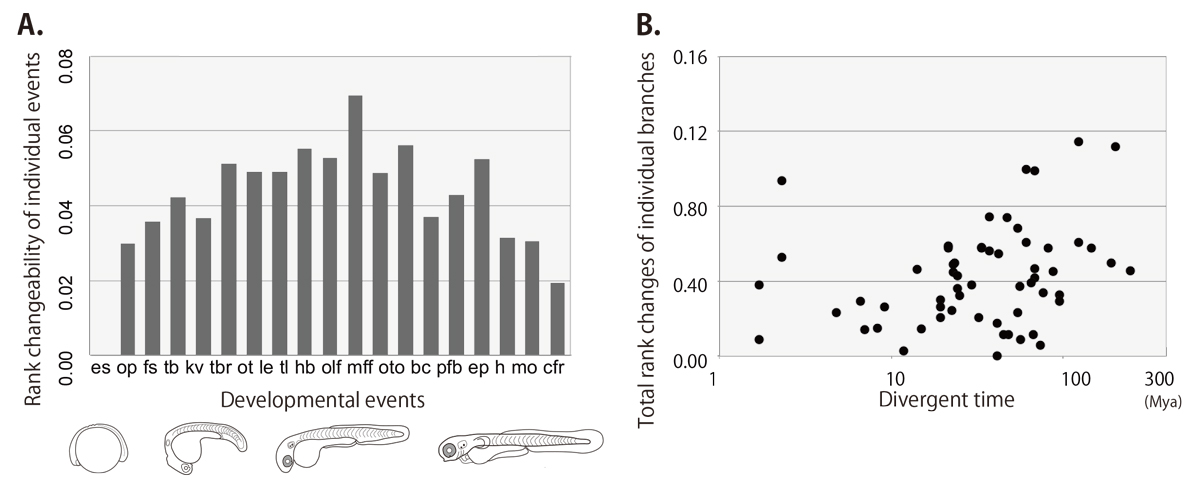
Figure: the rank changeability of individual developmental events (A) and the relationship between rank changes and divergent times (B) in teleost lineage estimated by the event-pairing method.










