



Is euchromatin really open in the cell?
Kazuhiro Maeshima*#, Shiori Iida*, Masa A. Shimazoe, Sachiko Tamura, Satoru Ide
*cofirst authors; #corresponding author
Trends in Cell Biology 2023 June 27 DOI:10.1016/j.tcb.2023.05.007
The human genome chromatin can be classified into euchromatin and heterochromatin, which have high and low transcription activities, respectively. In the classical view, it was believed that euchromatin has an open and decondensed structure, while heterochromatin is highly condensed.
A research team led by Professor Kazuhiro Maeshima of Genome Dynamics Laboratory (NIG), including a graduate student (JSPS Research Fellow DC2) Shiori Iida, SOKENDAI graduate student Masa A. Shimazoe, Technical Stuff Sachiko Tamura, and Assistant Professor Satoru Ide have put forward the model that euchromatin essentially forms condensed domains with sizes ranging from 100-300 nm in diameter based on new evidence from genomics and advanced imaging studies. They discuss this novel view of euchromatin in the cell and how the revealed organization is relevant to genome functions.
Physically condensed domains provide a higher-order regulation of transcription, while the extended fiber loops do not. Condensed domains likely hinder the accessibility of large transcription complexes to their target sequence located in the inner core of chromatin domains.
This exclusion effect can repress unintended gene expression. Although such a core of condensed domains seems to be inaccessible, condensed domains have the liquid-like property, allowing small transcription factors to penetrate the core of domains. Such small transcription factors may help a target sequence to float up to the domain surface to be transcribed. The authors suggest that condensed chromatin domains can work as Lego blocks of mitotic chromosomes during cell division, which create a smoother process for chromosome assembly and disassembly.
This work was supported by JSPS Fellowship, JSPD grants (JP21H02453, JP22H05606, JP21H02535, JP20H05936, JP16H06279(PAGS)), JSPS Research Fellow (JP23KJ0996(DC2)), JST SPRING (JPMJSP2104).
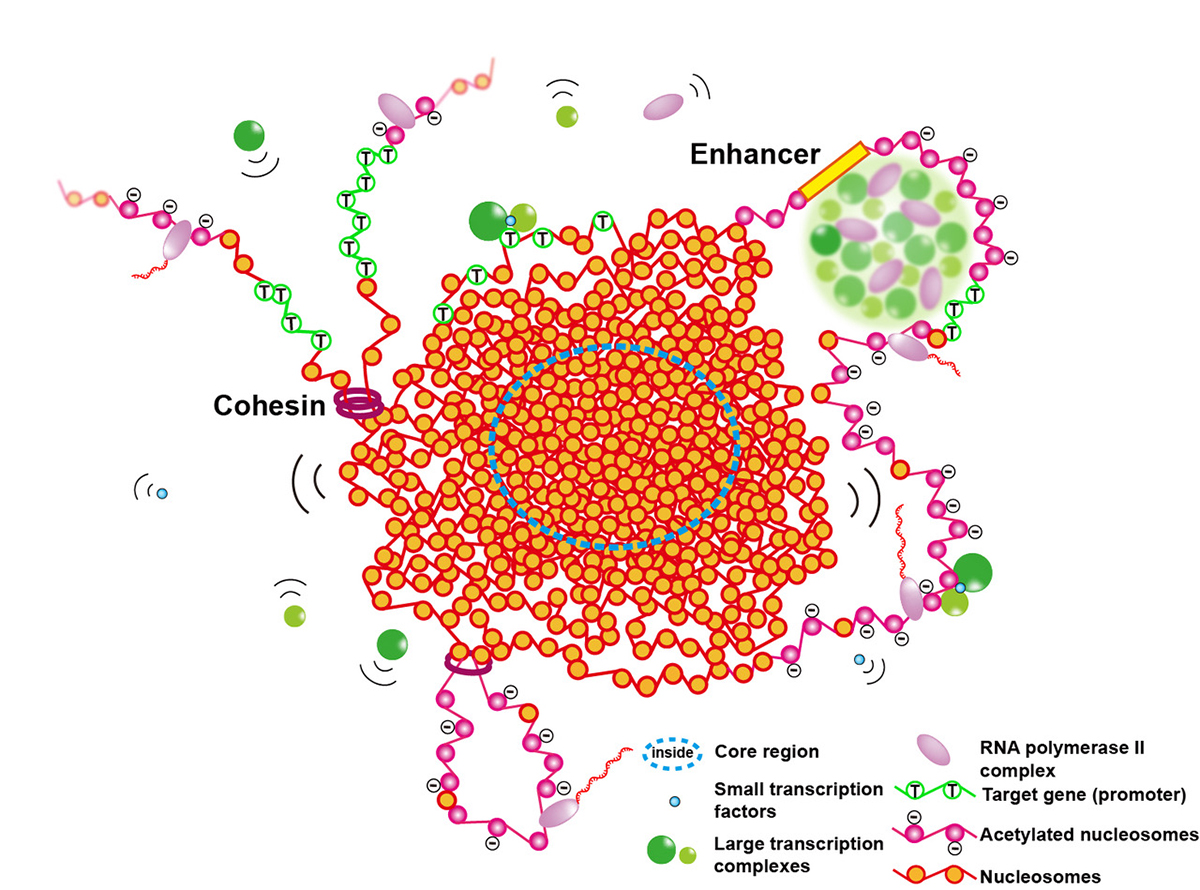
Figure: Euchromatic condensed domains provide higher-order regulation of transcription by physically excluding large transcription complexes (green spheres) from the inner core of the domains. Liquid-like property inside a condensed domain gives a certain degree of accessibility for gene expression. Small transcription factors interact with the target gene (open green circle nucleosomes) inside the inner core and cause it to float up to the domain surface to be transcribed. The condensed chromatin domains can work as Lego blocks of mitotic chromosomes during cell division.
第一次試験「東海・北陸地区国立大学法人等職員採用試験」
第一次試験 7月 2日(日)
第一次試験合格者発表 7月20日(木)9時30分
■機関説明会■
第一次試験に合格された方について、以下のとおりオンラインにて機関説明会を実施します。
(実 施 日) 7月27日(木)13:30~(30分程度)
(内 容) オンラインにて機関の紹介、質疑応答を行います。
(参加方法) ご希望者は、7月26日(水)正午までに、以下URLよりお申込みください。
https://forms.gle/yEDBvqtVPdCDekQe6
*返信にて、Zoom情報をお知らせいたしますが、届かない場合には、以下担当までご連絡願います。
第二次試験(1次選考・最終選考) ※ 新型コロナウイルス感染症の状況によっては、実施方法を
オンライン面接などに変更する場合があります。
■1次選考■
(対象者) 令和5年度東海・北陸地区国立大学法人等職員採用試験(事務、電気及び機械)の第一次試験合格者
(日 時) 令和5年8月7日(月) 予備日:9日(水)
(会 場) 情報・システム研究機構国立遺伝学研究所 本館2階応接室
(https://www.nig.ac.jp/nig/ja/about-nig/access_ja)
(実施方法) 個別面接
(事前に送付する物) ・訪問カード(履歴、志望動機等)
※所定の様式をダウンロードして記入の上、8月4日(金)10時までに、下記申込み先まで送付願います。
(持参する物) ・第一次試験合格通知のメール(写)
■最終選考■
(対象者) 1次選考の合格者
(日 時) 令和5年8月31日(木)から9月1日(金)
(会 場) 情報・システム研究機構本部事務局
東京都港区虎ノ門4-3-13ヒューリック神谷町ビル2階
(https://www.rois.ac.jp/about/access.html)
※最終選考は、情報・システム研究機構本部で実施します。
【 申 込 方 法 】
令和5年8月2日(水)15:00までに
「受験番号、氏名、メールアドレス、電話番号」を人事・労務係(
POLR1A variant- and tissue-specific effects underlie phenotypic heterogeneity in craniofacial, neural and cardiac anomalies
Kelly Smallwood, Kristin E.N. Watt, Satoru Ide, Kristina Baltrunaite, Chad Brunswick, Katherine Inskeep, Corrine Capannari, Margaret P. Ada, Amber Begtrup, Debora R. Bertola, Laurie Demmer, Erin Demo, Orrin Devinsky, Emily R. Gallagher, Maria J. Guillen Sacoto, Robert Jech, Boris Keren, Jennifer Kussmann, Roger Ladda, Lisa A. Lansdon, Sebastian Lunke, Anne Mardy, Kirsty McWalters, Richard Person, Laura Raiti, Noriko Saitoh, Carol J. Saunders, Rhonda Schnur, Matej Skorvanek, Susan L. Sell, Anne Slavotinek, Bonnie R. Sullivan, Zornitza Stark, Joseph D. Symonds, Tara Wenger, Sacha Weber, Sandra Whalen, Susan M. White, Juliane Winkelmann, Michael Zech, Shimriet Zeidler, Kazuhiro Maeshima, Rolf W. Stottmann, Paul A Trainor, K. Nicole Weaver
The American Journal of Human Genetics (2023) 110, 809-825 DOI:10.1016/j.ajhg.2023.03.014
Heterozygous pathogenic variants in POLR1A, which encodes the largest subunit of RNA Polymerase I, were previously identified as the cause of Acrofacial Dysostosis, Cincinnati-type. The predominant phenotypes observed in the initial cohort of 3 individuals were craniofacial anomalies reminiscent of Treacher Collins syndrome. We subsequently identified 17 additional individuals with 12 unique (11 novel) heterozygous variants in POLR1A and observed numerous additional phenotypes including neurodevelopmental abnormalities and structural cardiac defects, in combination with highly prevalent craniofacial anomalies and variable limb defects. To understand the pathogenesis of this pleiotropy, we modeled an allelic series of POLR1A variants in vitro and in vivo. In vitro assessments demonstrate variable effects of individual pathogenic variants on ribosomal RNA synthesis and nucleolar morphology which supports the possibility of variant-specific phenotypic effects in patients. To further explore variant-specific effects in vivo, we used CRISPR/Cas9 gene editing to recapitulate two human variants in mice. Additionally, spatiotemporal requirements for Polr1a in developmental lineages contributing to congenital anomalies in patients were examined via conditional mutagenesis in neural crest cells (face and heart), the second heart field (cardiac outflow tract and right ventricle), and forebrain precursors in mice (Figure A). Consistent with its ubiquitous role in the essential function of ribosome biogenesis, we observed that loss of Polr1a in any of these lineages causes death of the affected cells and resulting embryonic malformations. Altogether, our work greatly expands the phenotype of human POLR1A-related disorders and demonstrates variant-specific effects and differential tissue-specific requirements that provide novel insights into the underlying pathogenesis of ribosomopathies.
Assistant Professor Satoru Ide and Professor Kazuhiro Maeshima of Genome Dynamics Laboratory performed in vitro assessment of POLR1A variants, which impact rRNA transcription and the nucleolar morphology in human culture cells expressing wild-type or mutant POLR1A. Using a machine learning algorithm called “wndchrm”, Ide and Maeshima found that nucleolar morphologies of the mutants (based on POLR1A foci) are somehow corelated with their rRNA transcription capacities or status (Figure B).
The part to which NIG contributed was supported by the Japan Society for the Promotion of Science (JSPS) grants (22H05606, 21H02535 to SI; 19H05273 ,20H05936 to KM) etc. The image analysis by the machine learning algorithm “wndchrm” was conducted in collaboration with Dr. Noriko Saito in The Cancer Institute of JFCR.
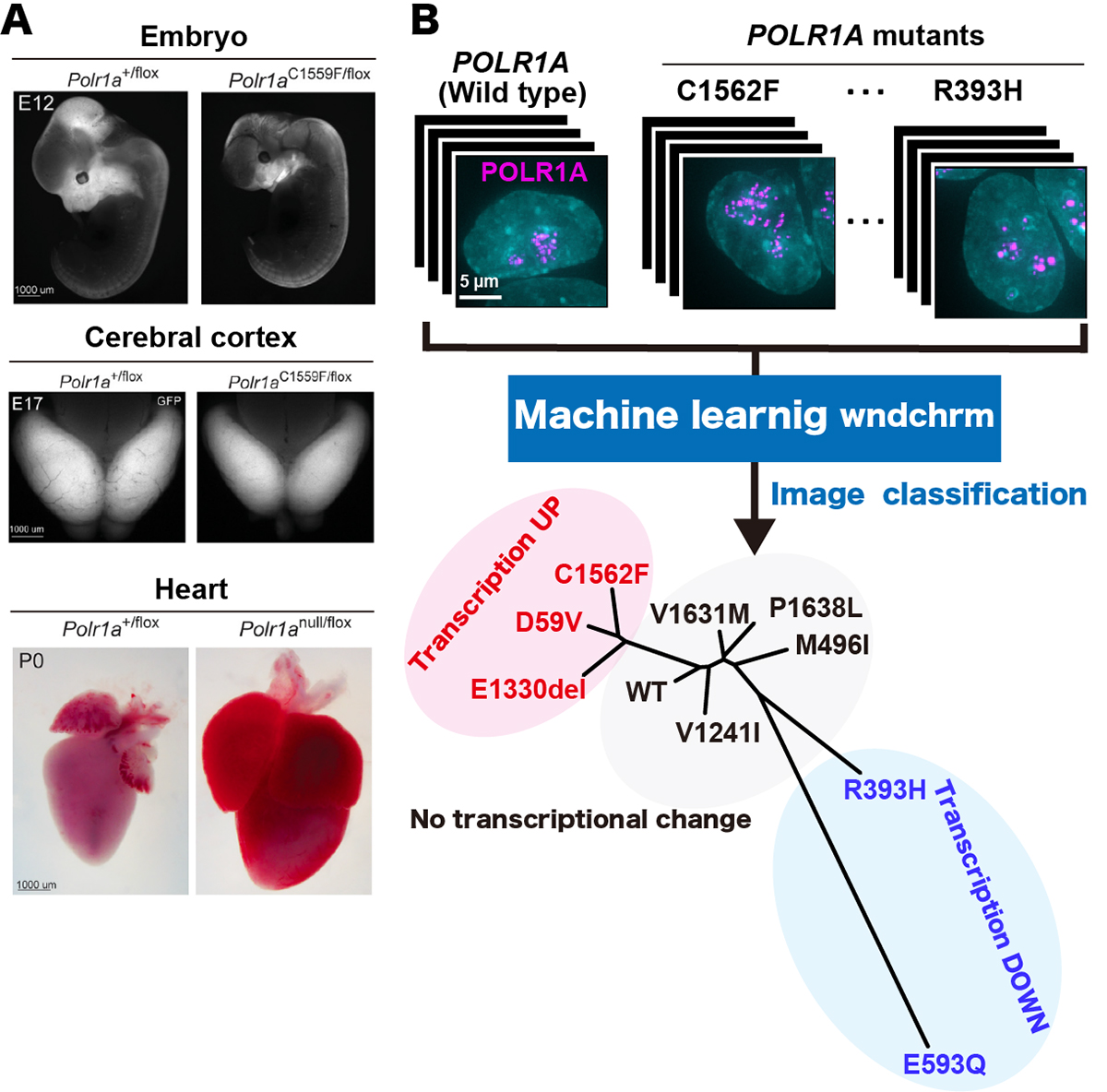
Figure: (A)Polr1a variant murine embryos demonstrate more hypoplastic craniofacial primordia at E12 (upper panel, right), a reduced cerebral cortex area at E12 (middle panel, right) and gross enlargement of heart at P0 (lower panel, right), compared to wild type (all the panels, left), respectively. (B) (upper panel) Localization of fluorescently labeled POLR1A (cellular nucleus, cyan and human POLR1A variants, magenta). (lower panel) Phylogeny shows the similarity of the nucleolar morphologies of wild type and nine POLR1A variants. POLR1A variants showing transcription upregulation, transcription downregulation and no transcriptional change are indicated in red, blue, and black, respectively.










