



Division-site localization of RodZ is required for efficient Z ring formation in Escherichia coli
Yusuke Yoshii, Hironori Niki, Daisuke Shiomi
Molecular Microbiology 2019 DOI:10.1111/mmi.14217
Cell elongation and cell division are controlled by different complexes. The mechanisms that promotes a switch from elongation to division have been still unclear. It has been recently reported that a direct interaction between MreB actin, which regulates elongation, and FtsZ tubulin, which regulates division, promotes the switching at the division site. We have analyzed RodZ that interacts with MreB. In this paper, we found that RodZ also localized on the division site depending on FtsZ, and that MreB localized to the division site depending on RodZ. When RodZ is not localized to the division site, the formation of the division ring (Z ring) composed of FtsZ was delayed. The localization of RodZ to the division site promotes the division ring formation and efficient cell division. This work was supported by KAKENHI and NIG-JOINT A.
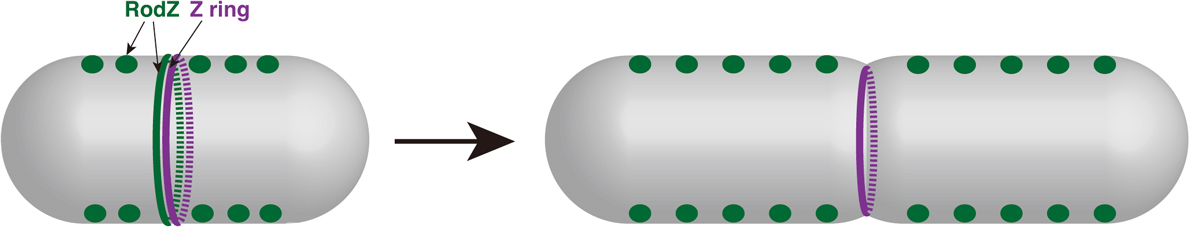
Figure: Division site localization of RodZ promotes an efficient formation of Z ring (i.e. switch from elongation to division).
Dynamic chromatin organization without the 30-nm fiber
Kazuhiro Maeshima, Satoru Ide and Michael Babokhov
Current Opinion in Cell Biology Volume 58, June 2019, Pages 95-104 DOI:10.1016/j.ceb.2019.02.003
Chromatin in eukaryotic cells is a negatively charged polymer composed of DNA, histones, and various associated proteins. Over the past ten years, our view of chromatin has shifted from a static regular structure to a dynamic and highly variable configuration. While the details are not fully understood yet, chromatin forms numerous compact domains that act as dynamic functional units of the genome in higher eukaryotes. By altering DNA accessibility, the dynamic nature of chromatin governs various genome functions including RNA transcription, DNA replication, and DNA repair/recombination. Based on new evidence coming from both genomics and imaging studies, we discuss the structural and dynamic aspects of chromatin and their biological relevance in the living cell.
This research was supported by JST CREST(JPMJCR15G2), JSPS Kakenhi (16H04746) and Takeda Science Foundation.
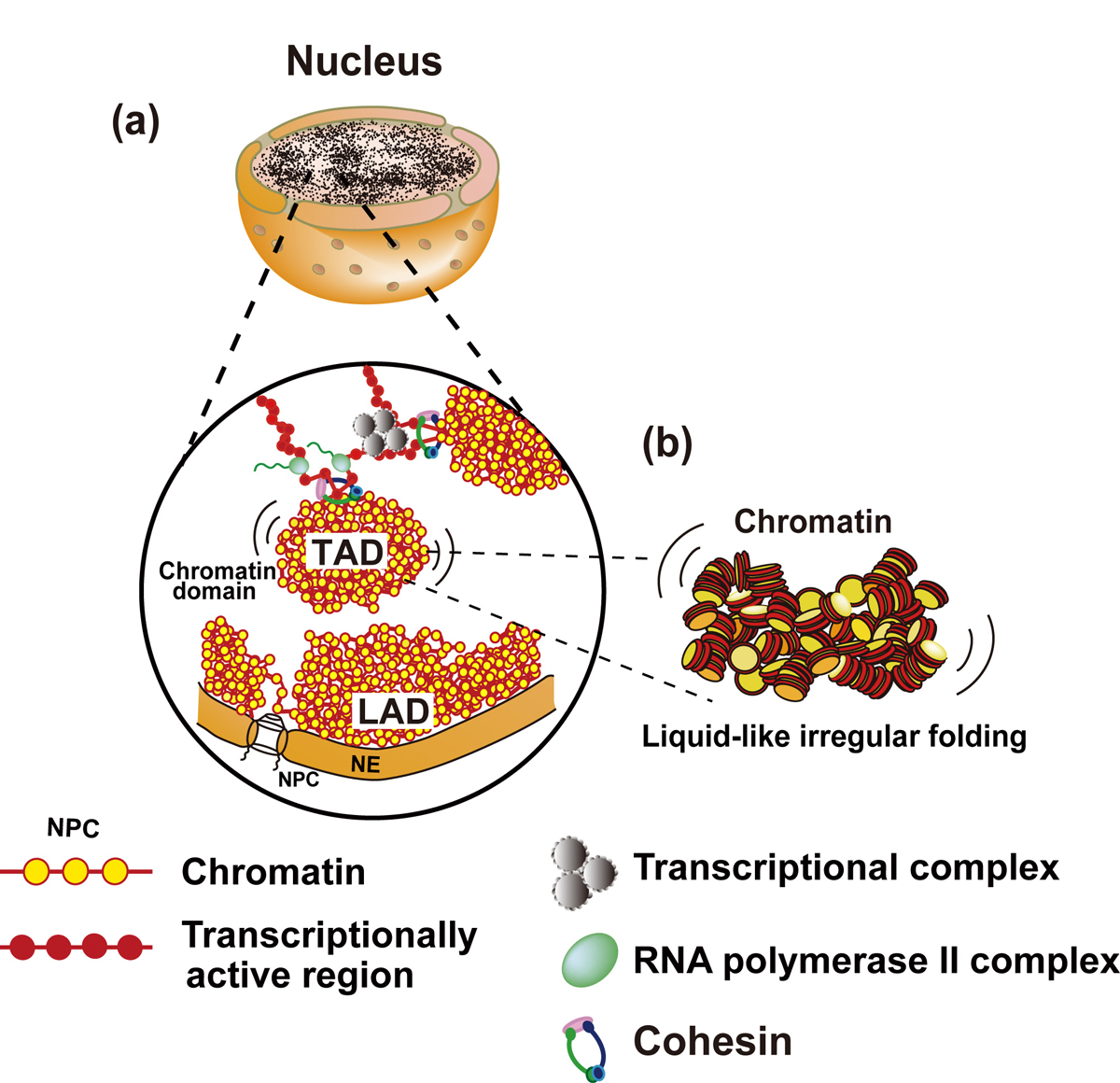
Figure: Interphase chromatin structure
(a and b) Chromatin consists of irregularly folded 10 nm fibers and forms numerous chromatin domains (e.g., topologically associating domains or contact/loop domains). The domains are formed by cohesin and nucleosome-nucleosome interactions. The compact domains could behave similar to a “liquid drop”. Binding of large transcriptional complexes (grey spheres) and RNA polymerase II (green spheres) might constrain movements of chromatin domains. Chromatin regions attached to nuclear envelop (NE) are called LADs (lamina-associated domains). NPC, nuclear pore complex; NE, nuclear envelop.
Press release
Single nucleosome imaging reveals loose genome chromatin networks via active RNA polymerase II
Ryosuke Nagashima, Kayo Hibino, S. S. Ashwin, Michael Babokhov, Shin Fujishiro, Ryosuke Imai, Tadasu Nozaki, Sachiko Tamura, Tomomi Tani, Hiroshi Kimura, Michael Shribak, Masato T.Kanemaki, Masaki Sasai, and Kazuhiro Maeshima
Journal of Cell Biology Published March 1, 2019 DOI:10.1083/jcb.201811090
Press release (In Japanese only)
The human body is composed of over forty trillion cells. Within each of these cells there is close to two meters of tightly packaged genomic DNA, the blueprint of life. Recently, there have been many advances in understanding how DNA is packaged and organized in the cell. In contrast, how DNA behaves mostly remains unknown.
In this recently published report, SOKENDAI graduate student Ryosuke Nagashima, assistant professor Kayo Hibino and professor Kazuhiro Maeshima of the National Institute of Genetics partnered with assistant professor S.S. Ashwin and professor Masaki Sasai of Nagoya University to study the movements of DNA in living cells using super resolution fluorescence microscopy (Figure 1A and B). Previously, it was commonly thought that the process of reading genomic information, known as transcription, would lead to more dynamic movements of DNA through the loosening of nucleosome packing. However, as Nagashima et al. report, transcription normally restricts DNA movement since the inhibition of transcription leads to increased DNA movements (Figure 1C). Furthermore, this work revealed that RNA polymerase II and other transcription-related factors form hubs to limit the motion of DNA (Figure 2). These results suggest that hubs may network the genome, restricting DNA movement to enable the efficient execution of transcription.
The results of this research have advanced our understanding as to how genetic information is acquired from DNA and may provide further clues concerning related disease states caused by abnormal changes to transcription.
This research is a result of a collaboration between the Genome Dynamics Laboratory of the National Institute of Genetics (Ryosuke Nagashima, Kayo Hibino, Michael Babokhov, Sachiko Tamura, Tadasu Nozaki, Ryosuke Imai, Kazuhiro Nagashima), the Nagoya University Department of Computational Science and Engineering (S.S. Ashwin, Shin Fujishiro, Masaki Sasai), the Cell Biology Center, Institute of Innovative Research, Tokyo Institute of Technology (Hiroshi Kimura), the Division of Molecular Cell Engineering of the National Institute of Genetics (Masato T. Kanemaki) and the Eugene Bell Center for Regenerative Biology and Tissue Engineering, Marine Biological Laboratory, Woods Hole (Michael Shribak, Tomomi Tani).
This research was supported by JST CREST(JPMJCR15G2), JSPS Kakenhi(16H04746), Takeda Science Foundation, RIKEN Pioneering Project, NIG-JOINT(2016-A2 (6)), SOKENDAI and National Institute of General Medical Sciences grant (R01-GM101701).
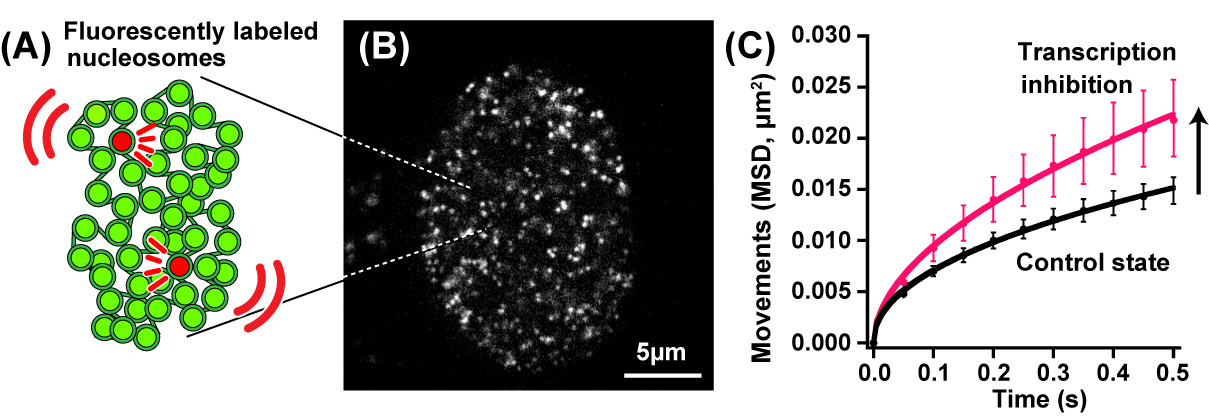
Fig.1. (A) A small fraction of nucleosomes was fluorescently labeled (red). (B) A single-nucleosomes image of a living human cell. (C) Genomic DNA motion, which is shown as mean square displacement, MSD), increased upon transcription inhibition.
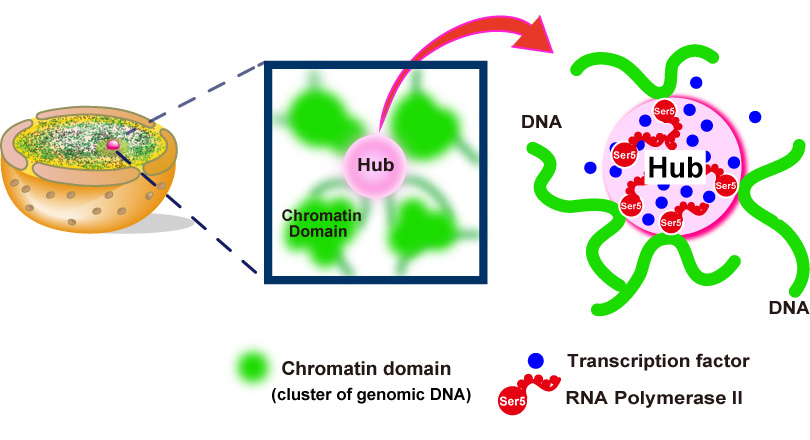
Fig.2. Hubs (pink spheres) of RNA polymerase II (red) and other components (blue spheres) of the gene transcription machinery constrain genomic DNA movements by connecting different genome regions together into an organized network.
Video1: Movie data (50 ms/frame) of single nucleosomes fluorescently labeled in a living human cell. Note that clear, well-separated dots and their movements were visualized.
Video2: Movie data (50 ms/frame) of single nucleosomes upon transcription inhibition. Note that greater movements of the dots are observed than those in Video1, suggesting the local genome dynamics increased upon transcription inhibition.










