



Molecular Function Laboratory • Kanemaki Group
Rapid Depletion of Budding Yeast Proteins via the Fusion of an Auxin-Inducible Degron (AID)
Kohei Nishimura and Masato T. Kanemaki Current Protocols in Cell Biology, 64, 20.9.1-20.9.16, 2014; DOI:10.1002/0471143030.cb2009s64Budding yeast has been an important eukaryotic model organism because there have been many genetic technologies available. In the yeast research field, temperature-sensitive (ts) mutants have been generated in order to inactivate a target protein conditionally. Although ts-mutants have been useful, there are problems as well; a drastic temperature shift causes heat-shock responses and there are some cases in which a target protein is not inactivated in a short period of time. To overcome these problems, we transplanted a plant-specific degradation pathway controlled by the plant hormone auxin to budding yeast and, in such yeast cells, a protein of interest can be degraded upon addition of auxin. We named this technology as the auxin-inducible degron (AID) system (Nishimura et al, Nature Methods, 2009) and subsequently improved it by making efficient degron cassettes (Kubota et al. Molecular Cell, 2013). The AID technology is getting popular in the filed of yeast studies. In this paper, we described a detailed protocol for generation of budding yeast AID mutants by one transformation. All plasmids and yeast strains required for the experiment are available from NBRP (http://yeast.lab.nig.ac.jp/nig/index_en.html). We hope that this protocol would help your study in the future.
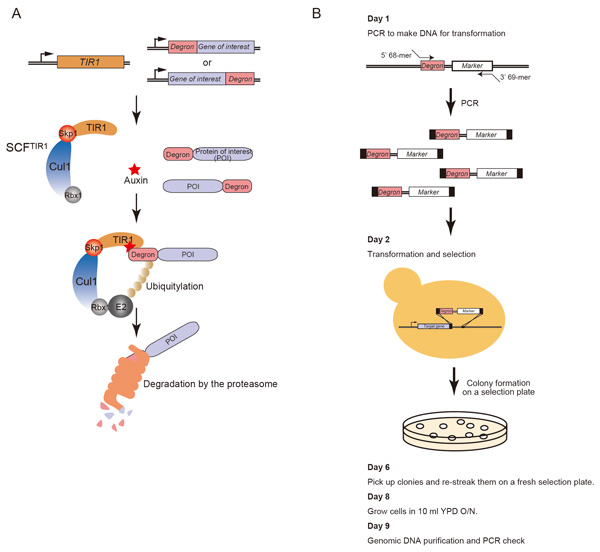
(A) Schematic illustration showing the mechanism of the auxin-dependent degradation of a degron-fused protein. (B) Schematic illustration showing the protocol for generation of AID mutants.
Division of Molecular Genetics • Fukagawa Group
The centromere: chromatin foundation for the kinetochore machinery
Tatsuo Fukagawa, and William C. Earnshaw Dev. Cell 30,496-508,2014; DOI:10.1016/j.devcel.2014.08.016Since discovery of the centromere-specific histone H3 variant CENP-A, centromeres have come to be defined as chromatin structures that establish the assembly site for the complex kinetochore machinery. In most organisms, centromere activity is defined epigenetically, rather than by specific DNA sequences. In this review, Fukagawa (Dep. Mol. Genet.) and Earnshaw (ex visiting professor of NIG, Edinburgh Univ) describe selected classic work and recent progress in studies of centromeric chromatin with a focus on vertebrates. We consider possible roles for repetitive DNA sequences found at most centromeres, chromatin factors and modifications that assemble and activate CENP-A chromatin for kinetochore assembly, and use of artificial chromosomes and kinetochores to study centromere function.
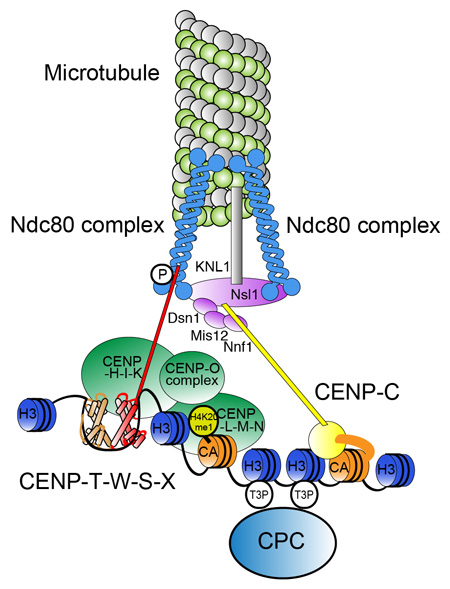
Molecular architecture of natural kinetochores. Centromere chromatin including is crucial for centromere specification and kinetochore assembly at natural centromeres. At the base of the structure are CENP-A containing nucleosomes, centromere specific H3 nucleosomes, and a CENP-T-W-S-X nucleosome-like structure in centromere chromatin. Centromere-specific chromatin structure is established by coordination of these components. CCAN proteins assemble on the centromeric chromatin and the microtubule-binding complex is subsequently recruited to assemble the functional kinetochore.










