



Microbial Genetics Laboratory・Niki Group
Comparative Genomics Laboratory・Fujiyama Group
Genome Biology Laboratory・Kohara Group
Wild-type E. coli are rod shape (Figure A, left). To make rod shaped cells, it is necessary that many proteins form complexes and function properly. E. coli cells are surrounded by rigid peptidoglycan (PG) layer and have to synthesize PG correctly. So far, we have identified RodZ required for determination of cell shape.
RodZ-deficient mutant is round shape (Figure A, middle) and grows slower than wild-type E. coli cell. To reveal function of RodZ protein, we isolated suppressor mutants that restore cell growth and shape of the rodZ mutant. To map the mutation sites in the suppressor mutants, we sequenced whole genome of twenty-nine mutants by a next-generation sequencer Solexa. This is the first report that mutation sites in ~30 mutants are determined by whole genome sequencing.
Most of the mutations were found in mreB, mrdA, or mrdB genes. It has been hypothesized that MreB, PBP2 encoded by mrdA gene, and RodA encoded by mrdB gene function with RodZ. Especially, twenty of twenty-nine mutants had a mutation in mreB gene. In addition, these mutations were clustered in domain 1A, one of the subdomain of MreB protein. These mutations change properties of MreB protein so that E. coli can form rod shape without RodZ protein. We also found that mutants of PBP2 and RodA change properties of MreB. Thus, we concluded that RodZ regulates function of MreB to form rod shape of E. coli.
This work was done by a collaboration of Niki lab, Fujiyama lab, and Kohara lab.
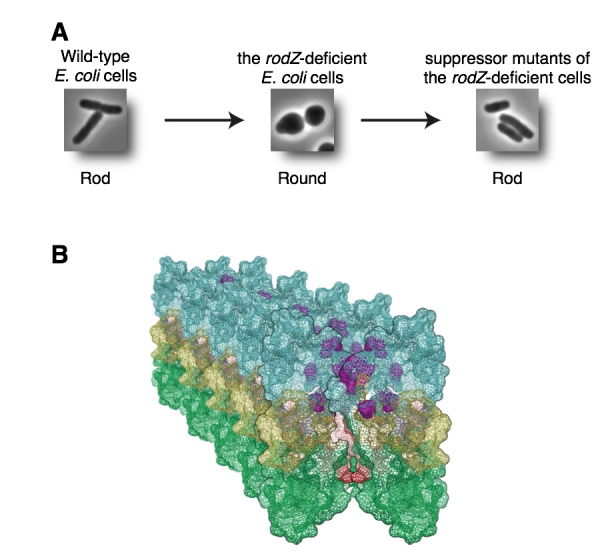
(A) Wild-type E. coli is rod (left). RodZ-deficient E. coli cell is round (middle). Suppressor mutants are rod (right). (B) Mutation sites shown by purple are clustered in domain 1A and it is a surface between MreB filaments. These mutations change properties of MreB filaments.
Division of Molecular Genetics・Fukagawa Group
The kinetochore forms a dynamic interface with microtubules from the mitotic spindle during mitosis. The Ndc80 complex acts as the key microtubule-binding complex at kinetochores. However, it is unclear how the Ndc80 complex associates with the inner kinetochore proteins that assemble upon centromeric chromatin. Here, based on a high-resolution structural analysis, we demonstrate that the N-terminal region of vertebrate CENP-T interacts with the “RWD” domain in the Spc24/25 portion of the Ndc80 complex. Phosphorylation of CENP-T strengthens a cryptic hydrophobic interaction between CENP-T and Spc25 resulting in a phospho-regulated interaction that occurs without direct recognition of the phosphorylated residue. The Ndc80 complex interacts with both CENP-T and the Mis12 complex, but we find that these interactions are mutually exclusive, supporting a model in which two distinct pathways target the Ndc80 complex to kinetochores. Our results provide a model for how the multiple protein complexes at kinetochores associate in a phospho-regulated manner.
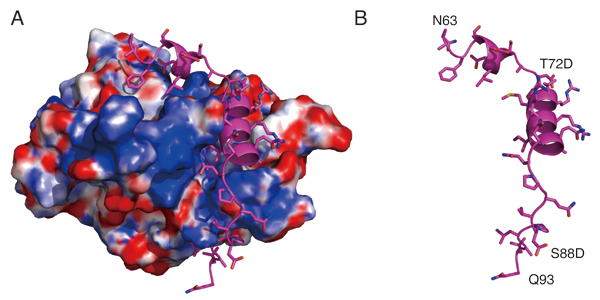
Structural model showing the surface charge of the Spc24/25 complex interacting with phospho-mimetic CENP-T peptide (Cyan). (B) Structural model showing the phospho-mimetic CENP-T peptide from the CENP-T-Spc24/25 complex structure in (A) on its own.

Mammalian Development Laboratory・Saga Group
In the mouse, testicular development is triggered in somatic cells by the function of Sry followed by the activation of fibroblast growth factor (FGF) 9, which regulates testicular differentiation in both somatic and germ cells. However, the mechanism is unknown. We show here that Nodal/Activin signaling pathway is activated in both male gem cells and somatic cells. The disruption of Nodal/Activin signaling drives male germ cells into meiosis and causes ectopic initiation of female-specific genes in somatic cells. Furthermore, we prove that Nodal/Activin-A works directly on male germ cells to induce male specific gene, Nanos2 independently of FGF9. We conclude that Nodal/Activin signaling is required for testicular development and propose a model in which Nodal/Activin-A acts downstream of FGF signaling to promote male germ cell fate and protect somatic cells from initiating female differentiation.
This study is conducted by Quan Wu who is a current student of SOKENDAI.
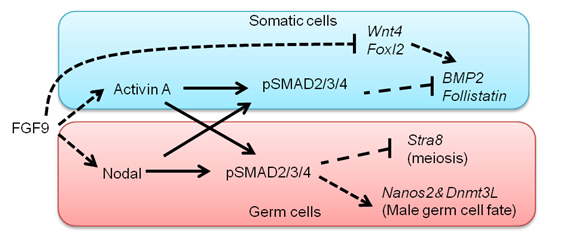
A model proposed by our study. FGF signals activate Nodal/Activin signaling pathway in both somatic cells and germ cells. Nodal/Activin-A triggers male sex differentiation by inducing male-specific genes, Nanos2 and Dnmt3L. Meanwhile, it suppresses Stra8 that is an essential gatekeeper of meiosis. In somatic cells Nodal/Activin-A thwarts the process of female differentiation by inhibiting Bmp2 and Follistatin.










