
生殖細胞の減数分裂に興味をもったきっかけは?
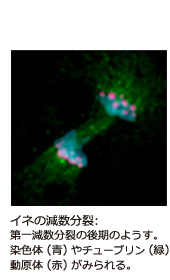
学部生時代に、先輩が顕微鏡でイネの染色体の減数分裂を観察しているのをのぞかせてもらい、非常に美しいと思ったことと、相同な染色体が対合するメカニズム に興味を覚えたのが、最初のきっかけです。減数分裂では、通常の細胞分裂の前に行われるDNA複製を挟まずに連続した2回の分裂がおき、遺伝情報を担う染 色体の数が親の半分になります。また1回目の分裂に先立ち、父親側と母親側の染色体が互いの相同性を認識して、同じ染色体同士がくっつき合う「対合」とい う現象がみられます。この過程により父親側と母親側の遺伝子がまざりあい、新しい遺伝子組み合わせをもつ精細胞と卵細胞が作り出され、子供へと引き継がれ ます。
私は農学部の育種学研究室に所属していたので、イネを素材に、実際に役立つ品種の育成をしたいと思っていました。結局は育種の道 には進みませんでしたが、 その後も一貫してイネを対象に研究を続け、14年前に遺伝研に赴任してからは、減数分裂やその前段階の過程についての本格的な研究を始めました。イネの変 異体の中から、花粉や種子ができないなどの表現型を選び出し、主に順遺伝学的な手法で機能解析を進めています。
今回、どのような遺伝子を同定し、減数分裂におけるどのようなことを明らかにされたのでしょう?
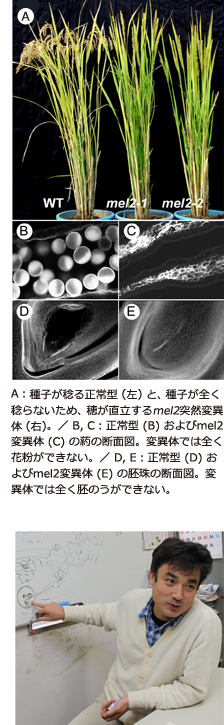
農業生物資源研究所で開発した突然変異系統のなかから、「種子が不稔となる突然変異体」を私たちのところで選び出して解析したところ、減数分裂への移行を制御するMEL2 というイネ遺伝子に行き当たり、その構造と機能を解明することができました。
被子植物が種子を残すためには、ある時期に一斉に雌雄の配偶体(すなわち花粉と胚のう)を作り出し、同じタイミングで受精を成立させるしくみが必要です。 たとえば、最初期にはたらく最も重要なしくみとして、日の長さや温度の変化を感知して一斉に生殖過程へと移行する現象が知られています。そのほかにもさま ざまなしくみがありますが、「減数分裂へ移行するタイミングの制御」もその一つです。葯の原基の中にたくさん作られる始原生殖細胞は、最初は各々がバラバラ(非同調的)に体細胞分裂を繰り返して細胞数を増やします。その後、ある時期に達すると、葯内の生殖細胞が同じタイミングで一斉に減数分裂に入ります。 被子植物の場合には、その後にさらに2回の体細胞分裂を経ることで花粉が完成します。同様に、めしべでも「減数分裂を雄側と同調させるしくみ」があると考えられます。しかし、減数分裂移行のしくみを支える植物の分子メカニズムは、これまで全くわかっていませんでした。
MEL2は、葯とめしべの中にある生殖細胞で特異的に発現していました。MEL2タ ンパク質のアミノ酸配列中には3つのモチーフがみられましたが、このモチーフ組み合わせをもつタンパク質は、今のところイネ科でしか知られていません。 モチーフの一つは、RNAとの結合が予測される「RNA認識モチーフ(RRM)」でした。このRRMは、ヒトのDAZAP1と、配列もモチーフもよく似て いました。DAZAP1は無精子症の原因タンパク質であるDAZと結合することが知られています。植物の減数分裂に関わる遺伝子がヒト無精子症の関連遺伝 子に似ていたことは、非常に興味深いと思います。
今回、私たちは、MEL2が体細胞分裂から減数分裂への移行のタイミングを制御するマスター遺伝子であると結論づけました。MEL2はイネ生殖細胞の細胞分裂周期、とくに「減数分裂の直前に特有のDNA複製の開始」を制御し、その制御システムが葯における減数分裂の同調性の確立とカップリングしていると考えています。そのため、mel2変異体では体細胞分裂から減数分裂へ移行できず、その結果として同調性も失われ、花粉が作られないと考えられます。mel2 変異体の葯の生殖細胞は、分裂を停止するか、あるいは体細胞分裂を繰り返し、やがてアポトーシスによって消滅してしまいます。
研究のどのような点が評価されたとお考えですか?
「花のでき方」を探る研究は国内外でさかんに行われており、遺伝子も多数同定されていますが、「植物の始原生殖細胞の誕生から減数分裂に至る過程」について は、これまで体系的な研究は行われていませんでした。そういう意味で、今回は「未知の領域に解明の糸口を見いだした点」と 「減数分裂への移行に必須の植物遺伝子を、世界ではじめて発見・解析した点」が評価されたと思っています。
といっても、研究は一筋縄ではいきませんでした。mel2変異体そのものは2000年にみつかりましたが、論文までには実に10年を要しました。生殖細胞の形成から種子の形成に至る「イネの一生」を追う必要があり、まさに時間と忍耐を必要とする研究でした。しかも、MEL2の ホモ変異体は種子ができないので、ヘテロ変異体を解析しなければならないという大変さもありました。未踏の領域だったので検体、すなわち「非常に幼い穂」 の収集・保存や解析手法の確立には、さまざまな試行錯誤が必要でした。結果として、イネの突然変異体を用いて、初期の生殖過程における細胞レベルでの事象 と個体レベルでの現象を結びつける独自性が生まれたと自負しています。
研究成果は農業などで応用可能なのでしょうか? また、今後の課題はなんでしょう?

すぐに農業に応用できるというものではありませんが、冷害などの環境ストレスによる種子不稔の対策に役立つ可能性があると考えています。たとえば、日本の東 北地方の太平洋側では「ヤマセ」という冷害が知られます。低温によってイネの生殖細胞を覆う細胞(タペート細胞)が肥大化して死んでしまい、減数分裂がで きずに花粉が作られなくなってしまうのです。mel2 変異体でもタペート細胞の肥大化がみられますので、減数分裂の進行阻害とタペート細胞の肥大化を解析することによって、イネの生殖細胞が高温や低温に弱い理由の解明やその対処法の検討に結びつくのではないかと考えています。
減数分裂への移行の制御、および減数分裂の同調性は、単子葉植物には共通してみられますが、今後は「シロイヌナズナなどの双子葉植物では、どの遺伝子がイネMEL2の機能を担うのか」についても調べていきたいと思っています。また、MEL2のRRMが結合するRNAの配列を早急に明らかにし、RNAと結合することで、MEL2が「どのような遺伝子発現機能を、どのように制御しているのか」を明らかにしたいと考えています。これまでに、MEL2を含めて5個の生殖関連イネ遺伝子の同定と解析を行ったことになりますが、全能性と永続性を合わせもつ生殖細胞がどのようにして作られるのかを、植物の一生との関わりにおいて明らかにしたいと考えています。
(文: サイエンスライター・西村尚子 / 2011.01.17掲載)
掲載論文
A novel RNA-recognition-motif protein is required for premeiotic G1/S-phase transition in rice (Oryza sativa L.)
Ken-Ichi Nonomura1,2,6, Mitsugu Eiguchi1, Mutsuko Nakano1, Kazuya Takashima1, Norio Komeda1,2, Satoshi Fukuchi2,3, Saori Miyazaki1,2, Akio Miyao4, Hirohiko Hirochika4, Nori Kurata2,5
- 国立遺伝学研究所・実験圃場
- 総合研究大学・生命科学研究科・遺伝学専攻
- 国立遺伝学研究所・遺伝情報分析研究室
- 農業生物資源研究所・基盤研究領域
- 国立遺伝学研究所・植物遺伝研究室
PLoS Genetics, 7(1): e1001265. DOI:10.1371/journal.pgen.1001265