遺伝子は染色体上の一定の座位に存在していて、染色体と行動をともにし、染色体の組換えや転座、欠失など染色体の構造的変異によってその座位を変えることがあっても、遺伝子が単独で、染色体から分離したり、他の染色体に移ることは一般的には考えられない。しかしアメリカのカーネギー研究所のMcClintock(1951)が、トウモロコシの種子や植物体に含まれるアントシアニン遺伝子や、胚乳の黄色デンプンの遺伝子などの発現を変化させる制御因子(control element)が、一つの染色体から他の染色体に移動して、その場所の遺伝子作用を調節することを見いだし、遺伝子が単独で、一つの染色体から他の染色体に移動することがあることがわかった。
このような遺伝子はトランスポゾン(動く遺伝子)といわれ、1960年代の後半に、大腸菌などの微生物でも発見され、転置因子(transposable element ; Tn)と呼ばれた。これら原核生物の転置因子は一定の塩基配列をもったDNAで、大腸菌などの正常の染色体の構成成分として1細胞あたり1~8個存在している。この因子は染色体に組込まれて存在するので挿入配列(insersion sequence ; IS)とも呼ばれ、分子の両端に16~38塩基対の反復した塩基配列をもっている。
「基礎遺伝学」(黒田行昭著:近代遺伝学の流れ)裳華房(1995)より転載
- 原核生物のトランスポゾン
このような転写因子はファージMuや性決定因子Fのようなプラスミドにも存在するし、種々の抗生物質抵抗性を支配するR因子にも存在する。このような原核生物のトランスポゾンを表1・4に示したが、R因子やMuの場合には、それ自体がそれぞれの遺伝情報をもっており、その長さも挿入因子よりも長く、200~20,000の塩基対からなり、その両端には800~1,500塩基対からなる反復配列(repeated sequence)をもっている。
表1・4:原核生物のトランスポゾン
名称 マーカー 分子の長さ(塩基対) 反復部の長さ(塩基対) Tn1 アンピシリン 4,957 38 Tn2 アンピシリン 4,957 38 Tn3 アンピシリン 4,957 38 Tn4 アンピシリン、ストレプトマイシン、スルフォマイド 20,500 短い Tn5 カナマイシン 5,400 1,500 Tn6 カナマイシン 4,200 電顕で検出できない Tn7 トリメソプリム、ストレプトマイシン 14,000 電顕で検出できない Tn9 クロラムフェニコール 2,638 18/23 Tn10 テトラサイクリン 9,300 11,400 Tn204 クロラムフェニコール、フシジック酸 2,457 18/23 Tn402 トリメソプリム 7,500 電顕で検出できない Tn501 水銀イオン 7,800 38 Tn551 エリスロマイシン 5,200 35 Tn554 エリスロマイシン、スペクチノマイシン 6,200 未決定 Tn732 ゲンタマイシン、トブラマイシン 11,000 未決定 Tn903 カナマイシン 3,100 1,050 Tn917 エリスロマイシン 5,100 短い Tn951 ラクトース 16,600 短い Tn1681 熱安定性のエンテロトキシン 2,088 768(IS1) Tn1721 テトラサイクリン 10,900 短い バクテリオファージ 38,000 なし Mu 「基礎遺伝学」(黒田行昭著;近代遺伝学の流れ)裳華房(1995)より転載
- ショウジョウバエのトランスポゾン
真核生物では上に述べたトウモロコシの例があるが、酵母でもチロシンのtRNAの抑制遺伝子supA tRNAtyrにもトランスポゾンが見いだされており、5,100塩基対のTy1の部分と、その両端に約300塩基対のデルタ(δ)の部分があり、酵母の半数染色体あたりTy1は35個、デルタは100個反復して存在していた。ショウジョウバエにおいては卵や培養した細胞に、数百から20,000の塩基対からなる種々の大きさの環状をした反復DNAが存在することが報告されている。このDNAの80%以上は中程度に反復したDNAで、ショウジョウバエの1ゲノムあたり100個近く含まれていて、その一つがコピア因子(copiaelement)といわれるものである。同様な反復DNAは他の研究者によっても発見され、約30種以上も見つかっているが、そのいくつかを表1・5に示した。
表1・5:ショウジョウバエのコピア様因子
分子量(キロ塩基対) LTR(塩基対) コピー数/ゲノム 標的(塩基対) 特異性 LTR両端の塩基配列 コピア 5.0 276 20~60 5 非特異的 TG……CA 412 7.0 481/571 40 4 非特異的 TG……CA mdg 1 7.2 442/444 20~30 4 非特異的 TG……CA B 104(roo) 8.5 429 80 5 非特異的 TG……CA mdg 3 5.5 267 15 (5) 非特異的 TG……AA ジプシー 7.3 ~500 5(0~25) ? ? ? BEL ~8 ? ~25 ? ? ? 297 7.2 414 30~50 4~5 (T)ATAT AGT……AC(T) 17.6 7.4 512 40 4~5 (T)ATAT AGT……AT(T) H.M.Sビーグル 7.3 266 50 4 ? AGT……ACT これらのコピア様因子は、5,000~8,500の塩基対から成り、いずれもその両端に260~570塩基対の長い端部反復配列(long terminal repeat ; LTR)をもっている。コピア因子は、宿主細胞のDNAから離脱すると、これがそのまま環状となって他の細胞に感染するほか、一度mRNAを転写し、これから逆転写酵素により多数のDNAを合成してこれが環状の状態で他の細胞に感染して、新しい細胞の中に入り、その細胞の染色体に組込まれることもある。この過程はニワトリの細胞に感染するレトロウイルスの増殖過程と類似しており、実際コピア因子とレトロウイルスの塩基配列を比較すると、転写のための信号としてホグネスボックスといわれる塩基配列やポリAが付加する塩基配列、その他両方のDNAには類似の構造がみられる。
ショウジョウバエのトランスポゾンにはコピア因子のほか、FB因子(fold back factor)といわれるものも10種近く見いだされており、宿主細胞の染色体からの離脱や挿入によって遺伝子の運び屋となったり、挿入箇所に不安定な突然変異を誘発することが知られている。
さらにまた、P因子(P factor)といわれるトランスポゾンもある。完全型のP因子は、それ自身2907塩基対のDNAからなりトランスポゼーズという染色体に挿入する際に必要な酵素のほか、抑制タンパク質を合成する遺伝情報をもっており、両端には31塩基対の反復配列をもっている。このP因子から片方の反復配列の23塩基対が欠損したヘルパーP因子(helper P factor)が生じたが、これはトランスポゼースを産生することができ、ヘルパー機能だけをもっている。また、完全型P因子よりトランスポゼーズの遺伝子を失ったベクターP因子(vector P factor)も見つかり、これは代わりに他の遺伝子を組込んで、それを細胞に運びこむことができる。
細胞の中に挿入した遺伝子DNAをベクターP因子に組込ませ、ヘルパーP因子とともにショウジョウバエの卵に微少注入すると、その卵の染色体DNAに新しい遺伝子を導入することもできる。ショウジョウバエのトランスポゾンであるコピア因子、FB因子、P因子を比較したものが図1・22である。
それぞれのトランスポゾンの基本構造の中で、両端に存在するLTR配列は、それぞれのトランスポゾンによって異なり、30~1,000塩基対である(図1・22A)。図1・22Bには、それぞれのトランスポゾン自身のDNAが示されており、コピア因子では3種のタンパク質の指令領域ORFI、ORFII、ORFIII、があり、FB因子では両端のLTR配列の中に、いくつかの短い塩基対が反復していることを示している。また、P因子にもタンパク質合成指令領域が存在している。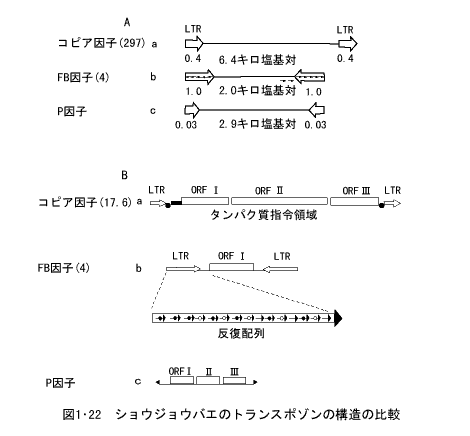
「基礎遺伝学」(黒田行昭著:近代遺伝学の流れ)裳華房(1995)より転載
- トランスポゾンの作用
以上述べた微生物や酵母、ショウジョウバエなどで見いだされたトランスポゾンは、一つの細胞の染色体から他の染色体に動くことによって、種々の遺伝的変化を生じることもわかってきた。図1・23に、これらのトランスポゾンが染色体を離脱し、他の染色体に移動して挿入されることによって惹起される変化を示した。トランスポゾンが単に挿入されるだけで、挿入された遺伝子は活性を失ったり変調を来すこともある。また染色体の一部が失われたり、欠失、逆位、転移などの遺伝子の組換えを生じる。
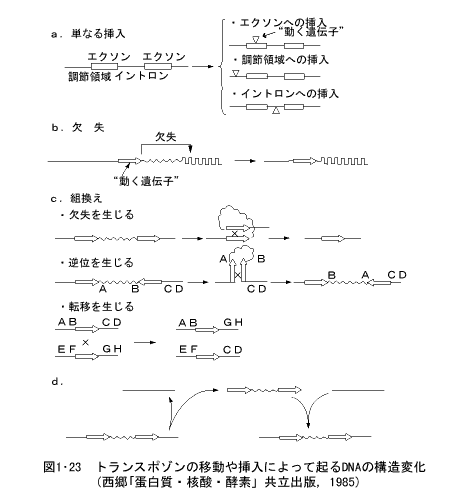
「基礎遺伝学」(黒田行昭著:近代遺伝学の流れ)裳華房(1995)より転載
